MolPlay: Democratizing Interactive Molecular Simulations and Analyses with a Portable, Turnkey Platform

M. Baaden, J. Phys. Chem. B 2024 https://doi.org/10.1021/acs.jpcb.4c04712 Computer-based tools for visualizing and manipulating molecular structures in real-time hold immense potential for accelerating research and improving education, but are only used to a limited extent. This paper explores the possibilities of these powerful techniques. It presents a classification of common interactive modeling tasks, such as […]
Introduction
The self-organisation of macromolecules into dynamic and hierarchical supra-molecular assemblies is a key feature of biological systems. Among those systems, soft structures, made of flexible polysaccharides, play a key role in the formation of biological hydrogels, as well as in the constitution of cellular membranes as extracellular/pericellular matrix. Even though pericellular coats play a crucial […]
Quartz Crystal Microbalance with Dissipation Monitoring (QCM-D)
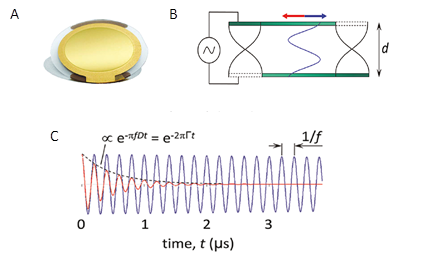
The QCM-D technique is widely used to study soft and solvated interfaces, because it enables both quantification of adsorbed mass and characterization of the viscoelastic properties of the adsorbed layer Reviakine et al., 2011. This techniques has found many applications to study fundamental biological and biotechnological questions as : adsorption of the proteins to the solid supports Muratsugu […]
Spectroscopic Ellipsometry
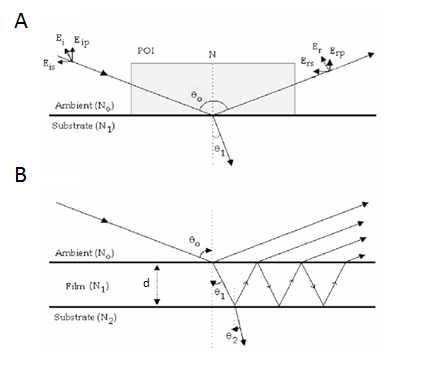
Ellipsometry is an optical technique that can be used to measure the mass, thickness and refractive index of adsorbed layers. In the frame of this thesis, ellipsometry was used as the main method to quantify surface densities of end-grafted HA films and incorporated proteins, as well as binding affinities and kinetics of HA-protein interactions. Working […]
Colloidal Probe Reflection Interference Contrast Microscopy (RICM)
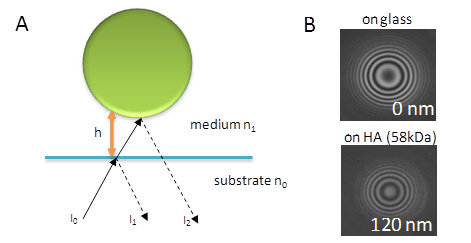
The RICM technique was first introduced to study cell adhesion, in particular for the visualization of the area of the cell attached to the surface in the focal adhesion site Curtis, 1964. In the 1980s, Sackmann et al. demonstrated that the distances between a planar transparent substrate and spherical objects, like a colloidal bead, hovering over […]
References
The Authors
Natalia S. Baranova Natalia graduated as a MSc in biochemistry from the Kiev National University in 2007. During her bachelor and master she has grown her interest in glycobiology. In 2008 she joined the group of Dr Ralf Richter (CICbiomaGUNE, San Sebastian, Spain) for a PhD project focused on the reconstitution of hyaluronan-rich extracellular matrix […]
Introduction
Many eukaryotic cells surround themselves with very soft and hydrated coats that are rich in polysaccharide matrix. Such coats play a fundamental role in regulating the interaction of cells with their environment. A crucial component of the coats is hyaluronan (HA), a regular, linear polysaccharide of the glycosaminoglycan family of typically several micrometers in contour […]
Biological Relevance of HA-Rich Matrices
In tissues, cells are embedded in extracellular matrix (ECM) which is made from components that are produced and then secreted by the cells. This matrix has many functions : it serves as integrative support to anchor cells, it distinguishes tissues from one another, it regulates inter-cellular communication and it ensures the mechanical stability of the tissue Alberts […]
The Cumulus Cell-Oocyte Complex Matrix
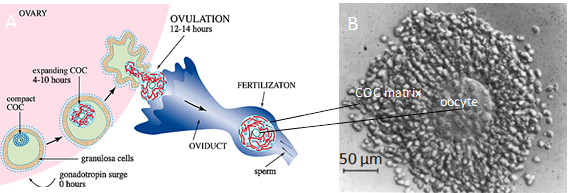
An illustrative example of a cross-linked PCM is the cumulus cell-oocyte complex (COC) matrix. This extended viscoelastic coat forms around oocytes just a few hours before ovulation and surrounds the oocyte on its journey towards fertilization in the oviduct. Before hormonal stimulation, the ovulating follicle represents a cavity filled with follicular fluid, where the wall […]
Composition of the Cumulus Cell-Oocyte Complex Matrix
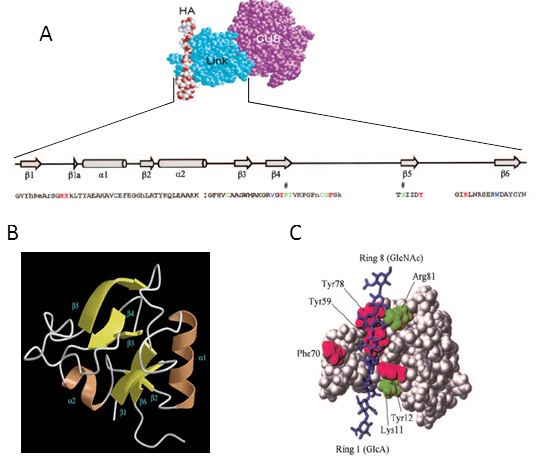
As outlined above, much of what is known about the mechanism of HA cross-linking comes from studies on the expansion of cumulus matrix. In particular, it was demonstrated that the presence of Hyaluronan(HA), Tumor Necrosis Factor-Stimulated Gene-6 (TSG-6), Inter-α-inhibitor (IαI) and Long Pentraxin 3 (PTX3) is essential for COC matrix expansion and stabilization Zhuo et al., 2001 Salustri et al. 2004Scarchilli et al., 2007 […]
Spatial and Temporal Regulation of COC Matrix Assembly
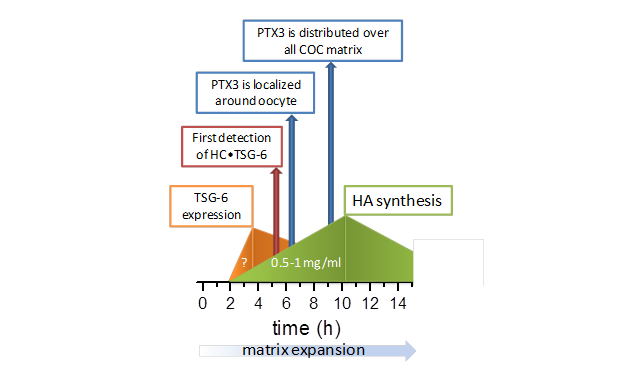
COC matrix formation is an illustrative case of spatio-temporal regulation of self-assembly. Because oocyte and cumulus cells are located in a special cavity, there is a spatial distribution of the crucial proteins : IαI is circulating in the blood and diffuses into the ovarian follicle only after initiation of ovulation, when the permeability barrier becomes leaky Powers […]
Model System to Reconstruct HA Matrix Assembly
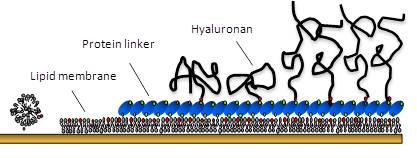
Biological surface science is an interdisplinary field which provides powerful tools to study biological phenomena. Well-defined biomimetic systems can be created thanks to numerous methods of surface fabrication. Information about the structural properties of surface-assembled ultrastructures can be obtained on the micro- and nanoscale by highly sensitive high resolution techniques Kasemo, 2002. A main scientific challenge […]
Conclusions and Perspective
Results gathered in a series of articlesBaranova et al., 2011Baranova et al., 2013Baranova et al., 2014provide novel mechanistic insights into the assembly and remodeling of HA-rich matrices under inflammation and inflammation like conditions. The quantitative data from the model systems demonstrate that full length TSG-6 forms oligomers upon interaction with HA which act as effective […]
References
The Authors
Natalia S. Baranova Natalia graduated as a MSc in biochemistry from the Kiev National University in 2007. During her bachelor and master she has grown her interest in glycobiology. In 2008 she joined the group of Dr Ralf Richter (CICbiomaGUNE, San Sebastian, Spain) for a PhD project focused on the reconstitution of hyaluronan-rich extracellular matrix […]
Introduction
Lignin (from the Latin lignum, meaning wood) is a major component of the vascular plant cell walls. Recently, the presence of lignin has been reported in nonvascular plants and red algae Martone et al., 2009 (Espiñeira et al., 2011). As one of the most abundant biopolymer on Earth, lignin is an amazing plant cell wall macromolecule in […]
Chemical Composition
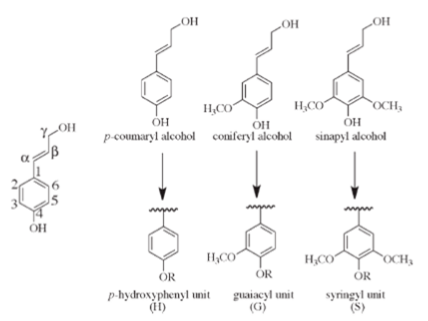
Conventionally, lignin is defined as a polyphenolic polymer basically made from three hydroxycinnamyl alcohols, called the monolignols, respectively, p-coumaryl, coniferyl and synapyl alcohols (Fig. 1), differing by the number of methoxyl ether groups on the ring. Other less represented monomer units originating from variations occurring into the monolignols biosynthesis, such as coumaryl, p-hydroxy-coumaryl etc…) may […]
Biosynthesis
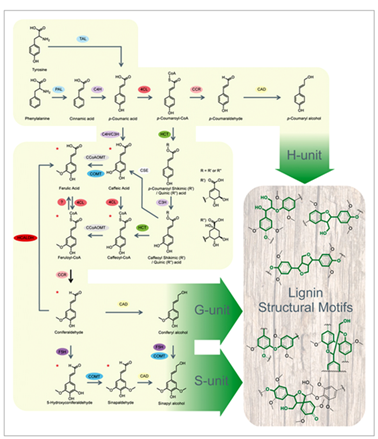
Four steps may be distinguished in lignin formation : 1/ biosynthesis of monolignols ; 2/ transport of monolignols to the plant cell wall ; 3/ enzymatic formation of monolignol radicals ; 4/ coupling of the radical monomers into the growing lignin polymer. In spite of progress in the understanding of lignin metabolism, parts of those are still controversial. Monolignol biosynthesisMonolignol biosynthesis derives from […]
Lignin Assembly

The chemical and biological mechanisms leading to lignin assembly in the plant cell wall into a complex 3D structure are not elucidated. Little is known about its supramolecular organization. What makes lignin polymerization in planta a peculiar phenomenon is that lignification consists of the formation of a hydrophobic polymer within the already organized chiral and hydrophilic polysaccharide […]
Cell Wall Lignification
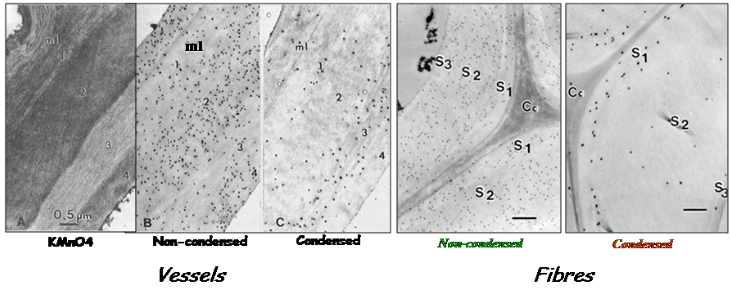
Biochemical studies supported these evidences of spatial heterogeneity of lignin deposition in cell walls associated to tissue maturation Joseleau et al., 1976. The genes involved in the spatio-temporal deposition of lignin are under the control of specific secondary wall NAC and MYB transcription factors Goicoechea et al., 2012 Nakano et al., 2015. In particular, MYB46/MYB83 directly activate MYB58/MYB63, […]
Lignin Valorization – A Challenge
Considering that lignocellulose biomass is one of the most abundant renewable resources, its comprehensive conversion into value-added products, including lignins, is of great importance due to its great abundance and its high carbon content. However, the difficulties of lignin extraction, its recalcitrance to common enzymatic fractionation together with its chemical variability makes of its valorization […]
References
The Authors
Katia Ruel Katia Ruel is actually retired from her position as Research Director at CNRS (Centre National de la Recherche Scientifique) where she pursued her scientific career at the Centre de Recherches sur les Macromolécules Végétales (CERMAV) in Grenoble (France). She received her education at the University of Orsay (Paris-XI) where she graduated in Biology. […]
Introduction
ABSTRACTThis review article aims at giving insight into cellulose-based nanomaterials and their potential use for wound dressing applications. The first part will focus on nanocellulose particles, including their production, properties and assembly into highly organized materials. The design of 2D (films and nanopapers) and 3D structures (cryogels and aerogels) will also be discussed, and the […]
From cellulose to nano crystals and nano fibrils
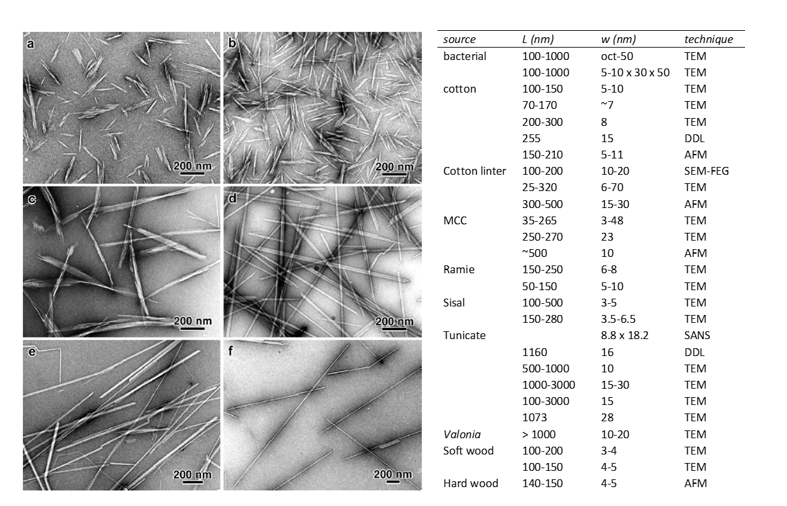
CELLULOSE NANOCRYSTALS (CNCs) Cellulose nanocrystals are rod-shaped particles of high crystallinity. They are isolated from the acid hydrolysis of the disorderd regions of the parent cellulose microfibrils, under controlled conditions of acid concentration, cellulose to acid ratio, temperature and time. Nickerson and Habrle first isolated them in 1947 (Nickerson and Habrle 1947) by cellulose degradation […]
Nano celllulose 2D dense structures
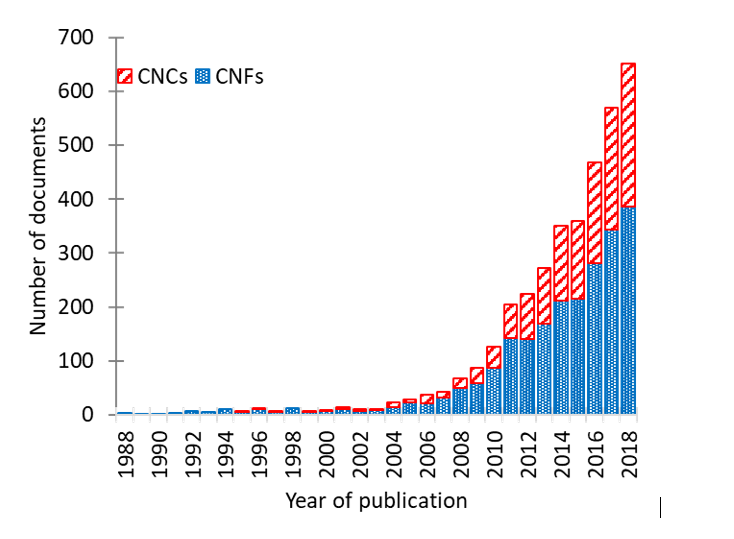
A wide range of nanocellulose particles, in term of morphologies and properties, can be obtained by playing on the preparations processes and the source of cellulose. From this diversity of cellulose nanoparticles, different solid structures can be prepared from various drying processes. The following sections deal with the production of nanocellulose 2D and 3D architectures, […]
Nano cellulose 3D porous structures
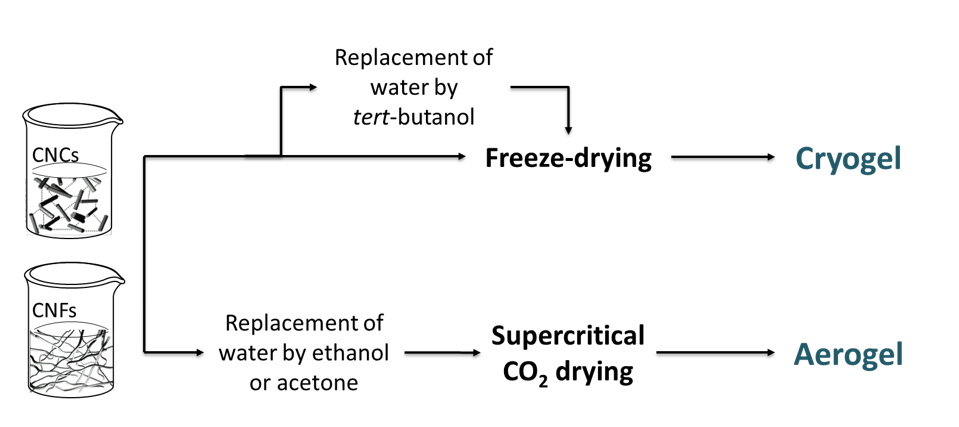
3D porous structures of nanocellulose There is an increasing wish for scaffolds in the biomedical, energy and thermal insulation fields. Nanocellulose 3D porous structures have been prepared to meet this need. In a recent review by De France et al. (2017), the preparation processes and material properties of CNC and CNF aerogels were described. A […]
Biomedical applications of nanocellulose 3-D materials
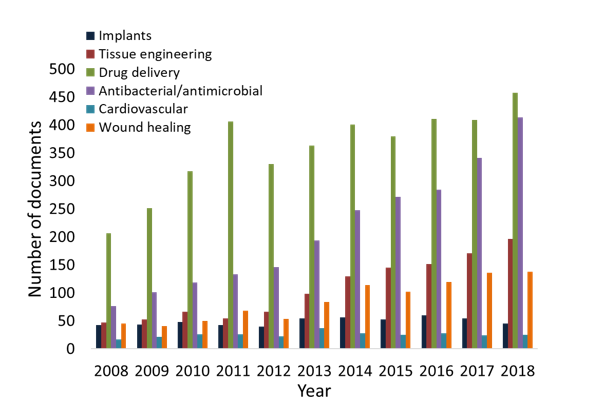
Porous nanocellulose materials have been prepared with various properties to meet the requirements for a wide range of applications, including the biomedical field. In the energy field, cryogels are used in solar cells as efficient and inert electrolyte deposition systems (Miettunen et al. 2014), and aerogels are used in the design of energy storage devices […]
Wound causing agents
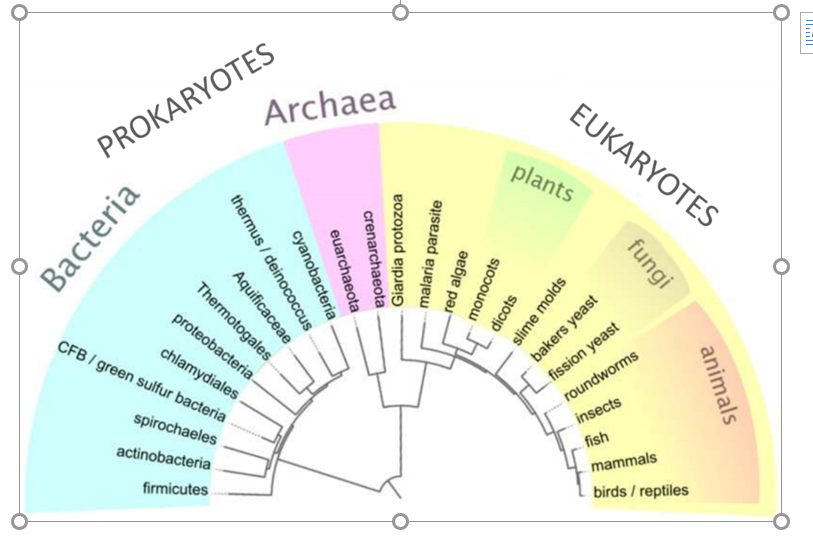
Microorganisms or microbes are defined as living microscopic organisms. They are found in very different parts of the phylogenetic tree (Figure 14) including bacteria, fungi, protozoa and algae. Figure 14: Phylogenetic tree of life of prokaryotes, covering both bacteria and archaea, and eukaryotes. The microorganisms represent most of life on Earth and are essential to […]
Wound infections
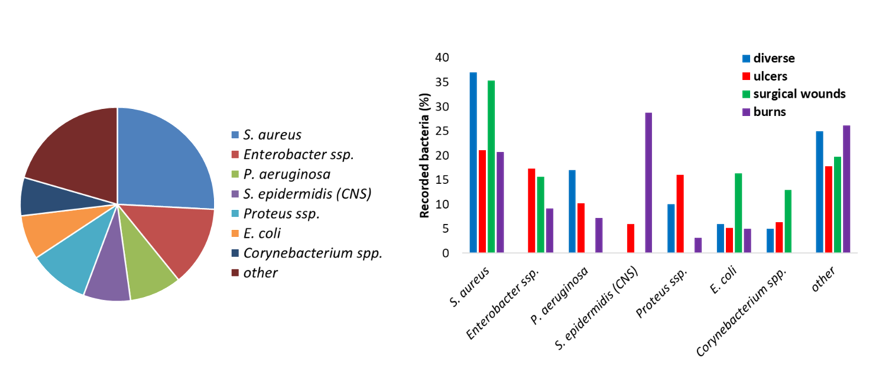
Wounds and wound healing process The skin is an organ protecting the human body from pathogens. A wound is a physical disruption of the skin, and therefore, its protective functions against the environment and pathogen microorganisms are lost. Wounds can be caused by abrasion, laceration, puncture, avulsion or burns. Chronic wounds, in contrast, are caused […]
Bioactive wound dressings
Bioactive wound dressings Cellulose: One of the advantages of cellulose in wound dressings is its ability to absorb water and therefore, exudate while providing a moist environment at the wound site (Basu et al. 2017). There are some commercially available cellulose-based dressings, such as Dermafill®, Nanoderm® and Vermac®. Bacterial cellulose, in particular, is used in […]
Nanocellulose functionalisation in traditional solvents
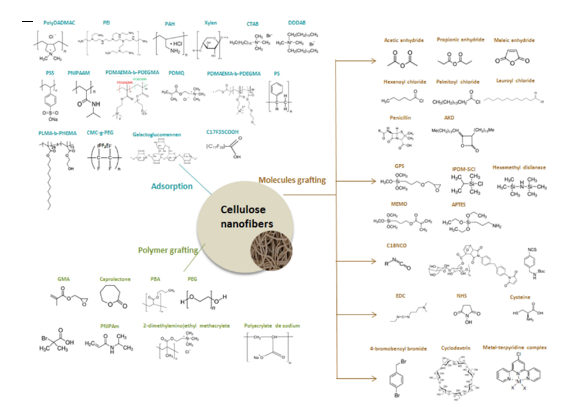
Nanocellulose present numerous interesting properties of renewability, non-toxicity, good mechanical properties, barrier properties, high specific surface areas etc. Applications areas seem to be endless, from packaging and paper industry to electronics, or even healthcare. To impart new functions to nanocellulose-based materials, the surface modification of CNFs or CNCs is sometimes required. Nanocellulose surface modifications have […]
Nanocellulose functionalisation in ‘green’ solvents
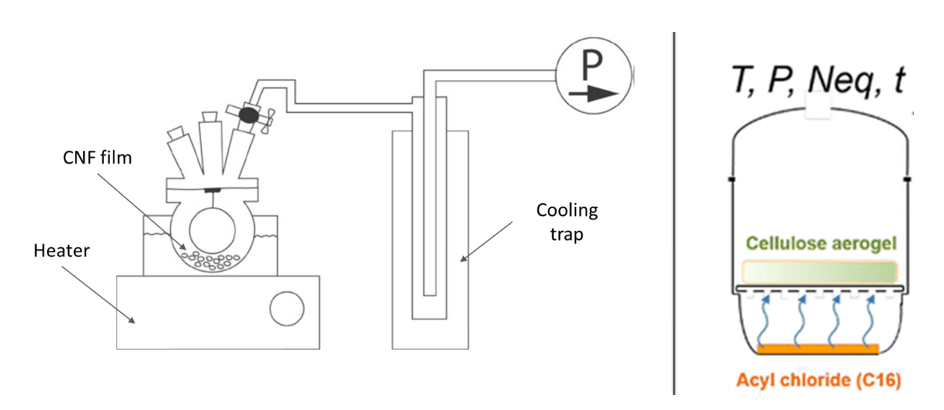
In the context of nanocellulose functionalization, the trend goes to greener processes to keep the interest of such sustainable materials. The development of safe and non-toxic modification strategies is essential to provide sustainable materials. The use of bio-based nanocellulose materials is by itself following the green chemistry principles. Green or sustainable chemistry implies the use […]
References
References
The authors
Clementine Darpentigny Clémentine Darpentigny studied polymers for advanced technologies at the Université Grenoble Alpes. During her PhD she investigated the use of supercritical carbon dioxide for the functionalization of nanocellulose-based structures. This work was performed under the supervision of Bruno Jean, Guillaume Nonglaton and Julien Bras and was a collaborative project between the Centre de […]
Introduction
The aim of this chapter was to give some representative, key examples in the domain of anticancer vaccines from the perspective of their design and assembly to their disclosed immunological data. Starting from the first historical approach, which is referred to as “semi-synthetic”, we will outline both its achievements and limitations ; alternatives for the antigen […]
Mono-TACA vaccines based on protein-carriers
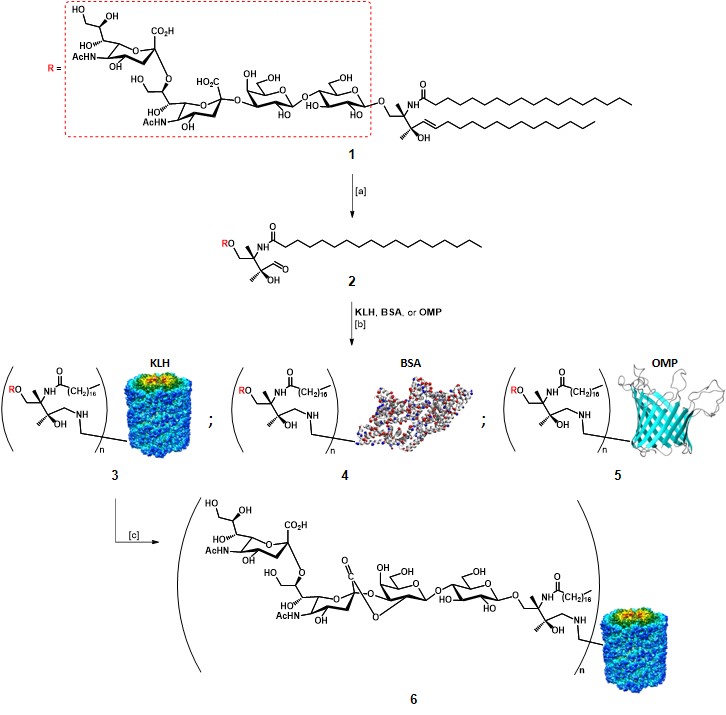
Mono-TACA vaccines based on protein-carriers As mentioned earlier, the first approaches for TACA-based vaccines relied on the utilization of extracted TACAs, in particular, GD2, GD3, and GM2.Slovin, Keding, & Ragupathi, 2005 Helling and co-workers first established that it was possible to stimulate an improved immune response in mice models by administration of a GD3 antigen covalently […]
Clustered mono-TACA vaccines based on protein-carriers
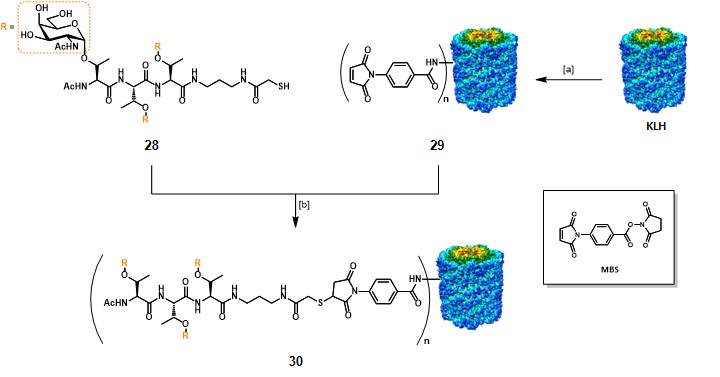
In the frame of clustered monomeric vaccines, Danishefsky’s research group reported the synthesis and immunological evaluations of a clustered Tn-based conjugate vaccine (30), where a cluster of three threonine-linked Tn antigens (28) was conjugated to the KLH carrier protein (Scheme 4).Kagan et al., 2005 The clustered B-cell epitope 28, containing a terminal sulfhydryl group was obtained […]
Hetero-TACA vaccines based on protein-carriers
Mono-TACA vaccines have shown promising results in early clinical trials, nevertheless this approach does not take account of the inherent heterogeneity of TACAs expressed on transformed cell surface. Danishefsky, Livingston, and co-workers hypothesize that the combination of different carbohydrate antigens, associated with a particular cancer type, could be beneficial for the induction of a more […]
Problems associated with protein carriers
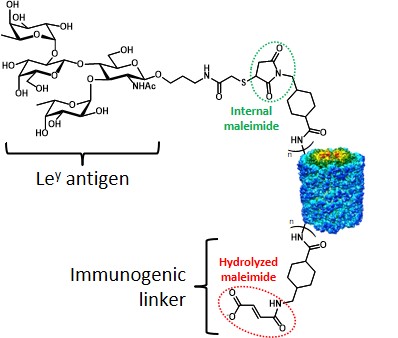
Despite these important results obtained in the field of chemical immunology, the carrier-protein approach suffers from some limitations. The TACA-protein conjugation chemistry represents a first, problematic aspect of the semi-synthetic strategy. In early models, as seen for GD3-protein conjugates, the sugar moiety underwent ozonolysis at the ceramide portion to give an aldehyde group which allowed […]
Alternatives to protein-carrier delivery
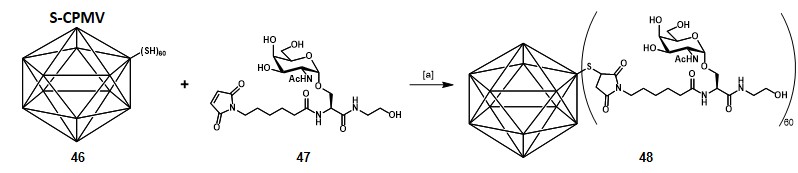
Virus-like particles Virus-like particles (VLPs) are non-infectious multiprotein structures which mimic envelopes or capsids of authentic native viruses, but lacking the viral genome.Roldão et al., 2010 They do not need attenuation or inactivation procedures, and their potency can be enhanced if immunosuppressive native viral-proteins are excluded from the VLPs composition. Their highly repetitive surface makes them […]
Two-component (TACA-CD4+), fully synthetic anti-cancer vaccines
Leclerc and co-workers paved the way for the fully-synthetic approach in TACA-based anticancer vaccines. In 1997, they prepared a glyco-dendrimer, namely multiple antigen glycopeptide (MAG), displaying the Tn antigen in its α-GalNAc-Ser form. (bay et al., 1997) Inspired by the multiple antigen peptide (MAP) described by Tam in 1988, (posnettz et al., 1988> the MAG […]
Two-component (TACA-lipopeptide), fully synthetic anti-cancer vaccines
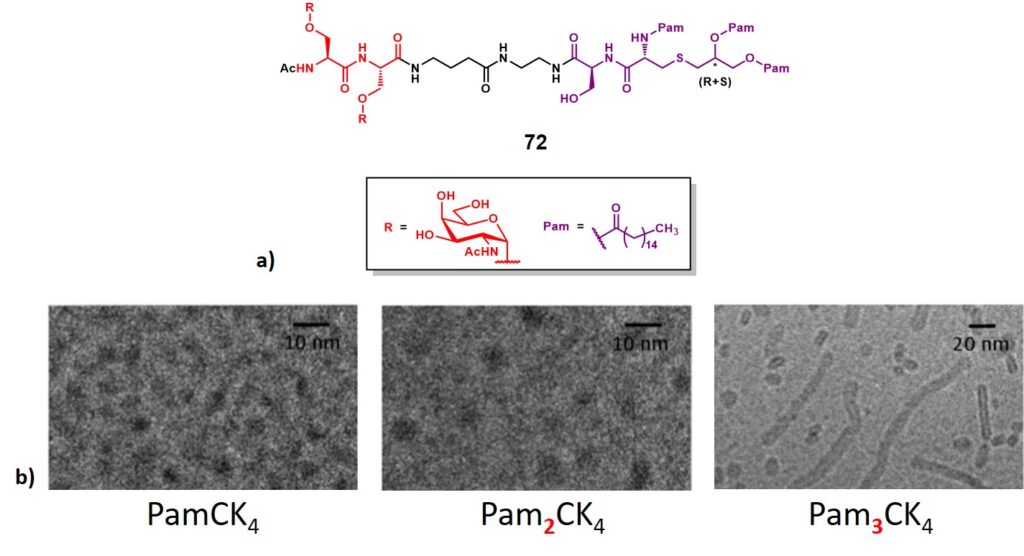
Along with the abovementioned two-component synthetic constructs bearing B- and TH-cell epitopes, other research groups reported a similar design strategy involving TACAs associated with a second module which differs from the CD4+ TH-cell epitope.Singhal and co-workers reported the synthesis and immunological evaluation of a dimeric (Ser)Tn-lipopeptide conjugate (72, figure 12a). This represents an early report […]
Multi-component, fully synthetic anticancer vaccines
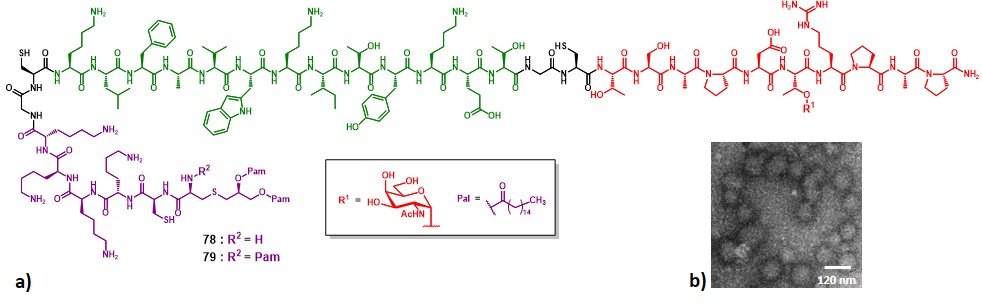
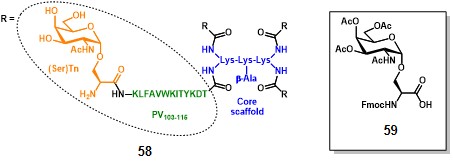
In 2007, Boons and co-workers described syntheses and immunological properties of three-component vaccine construct 78 and 79 (Figure 15a). These multicomponent constructs incorporate a glycopeptide MUC-1 sequence as B-cell epitope (in red), the PV103-115 class-II restricted TH epitope (in green), and the TLR-2/6 activator Pam2CK4 (78, R = H), or the TLR-1/2 activator Pam3CK4 (79, R = Pal) (in […]
Adjuvants
To complete this overview on vaccine design, it is noteworthy at least mentioning the wide role of adjuvants : immune-enhancing additives employed since the earliest attempts to raise significant immune responses. All adjuvant systems are basically focused on two mechanisms : (i) specific immune activation, and/or (ii) delivery-depot effect. Despite the abundance of adjuvant systems employed in […]
Conclusions
Semi-synthetic approaches In this section, a panel of strategies to overcome the problems associated to the semisynthetic protein-carrier approach has been presented. Still today, the lack of a fine TACA-protein conjugation control and the problems associated to the characterization of the resulting vaccine construct, together with other difficulties related to the inherent immunogenicity of protein […]
References
The author
Carlo Pifferi Carlo Pifferi obtained his Master’s degree in Pharmaceutical Chemistry and Technology, under the supervision of Prof. C. Nativi, Dr. S. Roelens, and Dr. O Francesconi at the University of Florence (Italy), focusing on supramolecular chemistry and organic synthesis of synthetic receptors for the molecular recognition of carbohydrates. He then attended the University of […]
Introduction
The essential cornerstones of cancer therapy have historically been surgery, radiotherapy, and chemotherapy. When the disease is identified early, and the tumor is small-sized and confined to a limited area, surgical resection of solid tumors still represents one of the most frequent local therapeutic choices. Nevertheless, large invasive tumors, most metastatic diseases, and hematological malignancies […]
Tumor-associated antigens
The importance of choosing the right antigen to develop anticancer vaccines represents a first, important challenge, which also prompted the understanding towards other key aspects. Following the achievements obtained by employing live attenuated pathogens vaccines, and because of the initial absence of defined tumor-associated antigens (TAAs), the first anticancer vaccines were constituted by whole tumor […]
MUC1 and tumor-associated carbohydrate antigens (TACAs)
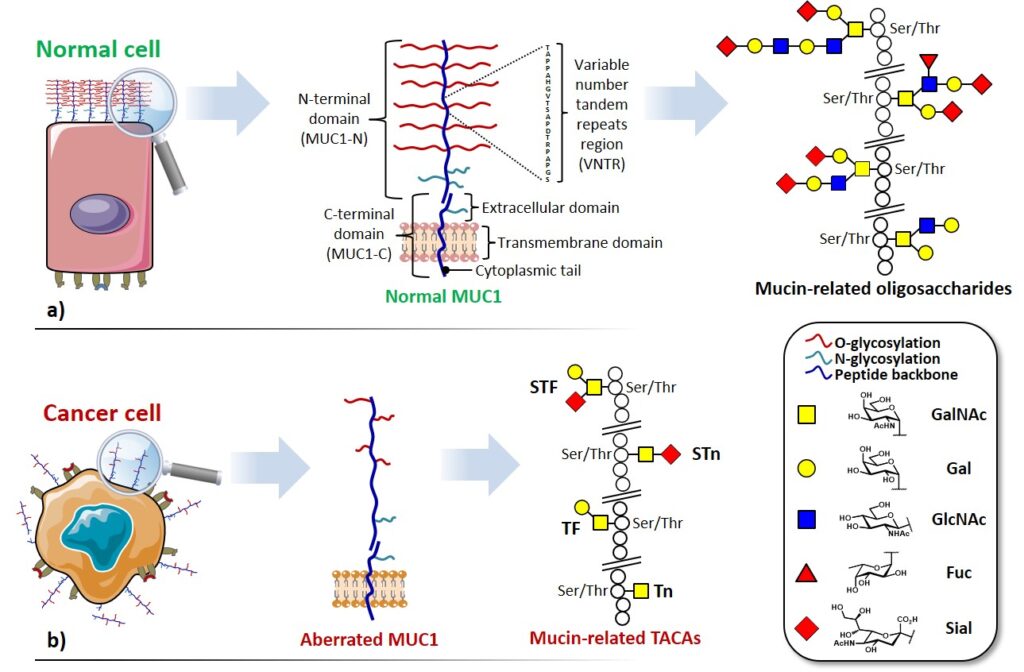
Among the abundance of shared tumor associated antigens, many research groups focused their interest on carbohydrate antigens related to the glycoprotein MUC1.Nath & Mukherjee, 2017 This heavily glycosylated transmembrane protein is composed of two fragments : (i) a longer extracellular N-terminal subunit (i.e. MUC1-N, 250-500 kDa), featuring a tandem repeat region (VNTR : variable number tandem repeat) of […]
Key aspects related to TACA-based vaccines development
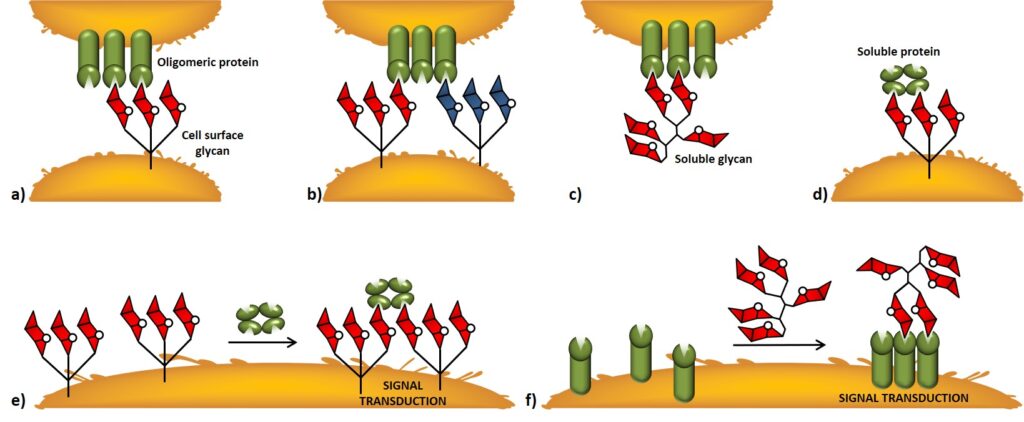
Source of epitopes The isolation of TACAs from natural material represents an extremely arduous task ; the first approaches in this field involved tedious and time-consuming operations to obtain the antigens of interest from highly heterogenic mixtures of sugar chains.Helling et al., 1994 The recent advances in glycobiology and synthetic organic chemistry allowed overcoming this primary aspect […]
References
The author
Carlo Pifferi Carlo Pifferi obtained his Master’s degree in Pharmaceutical Chemistry and Technology, under the supervision of Prof. C. Nativi, Dr. S. Roelens, and Dr. O Francesconi at the University of Florence (Italy), focusing on supramolecular chemistry and organic synthesis of synthetic receptors for the molecular recognition of carbohydrates. He then attended the University of […]
Abstract
Despite the recognition of the major importance of carbohydrates among all the natural disciplines, our present knowledge on their roles in modulating different processes of life is still limited. This is particularly true for the understanding of how carbohydrates trigger immunological functions. Typing “carbohydrate AND immunology” as a query on ISI Web of Knowledge lead to […]
Old and Hidden Works: A Chronology

1893. Defining Carbohydrates. “It appears, therefore, that we have an homologous series of aldehyde or ketone alcohols of the general formula CnH2nOn with these common properties: With this list, W. E. Stone described the use of the term “Carbohydrates” in 1893. w.e. stone, 1893 The term “carbohydrates” was well defined and explained to the scientific […]
Conclusions
All together, these works represent a clear rational journey in the field of glycoscience. Some of the described works are “hidden” in literature but still full of valid conclusions and insights. This chapter was designed with the idea to stimulate young and experienced glycoscientists with old seminal papers. From the definition of the term “carbohydrates” till […]
References
• Avery O. T. and Goebel W. F., Chemo-immunological studies on conjugated carbohydrate-proteins. J Exp Med, 1931, 54(3) : p 437-47 • Boyd W. C., Lectins. Annals New York Academy of Sciences, 1970, p. 168-90 • Clegg J. A. and Smithers S. R., Acquisition of Human Antigens by Schistosoma mansoni during cultivation in vitro. Nature, 1971, 232, p. 653-4 • Fishman P. H. and […]
The Author
Fabrizio Chiodo Fabrizio studied at the University of Palermo (Italy) where he got his Master degree in Pharmaceutical Sciences in 2008. At the end of 2008 he stared his PhD in the group of S. Penades (CIC BiomaGUNE, San Sebastian, Spain) under the supervision of M. Marradi. He was involved in different projects dealing with carbohydrates […]
Introduction
Sialic acid-binding immunoglobulin-like lectins (Siglecs) are a family of transmembrane proteins able to recognize sialic acids, nine-carbon atoms sugars highly expressed on the cell surface and one of the major components of the glycocalix. Sialic acids usually occur at the terminal positions of oligosaccharides and glycoconjugates, and they can act as ligands for receptor recognition […]
The Siglec Family
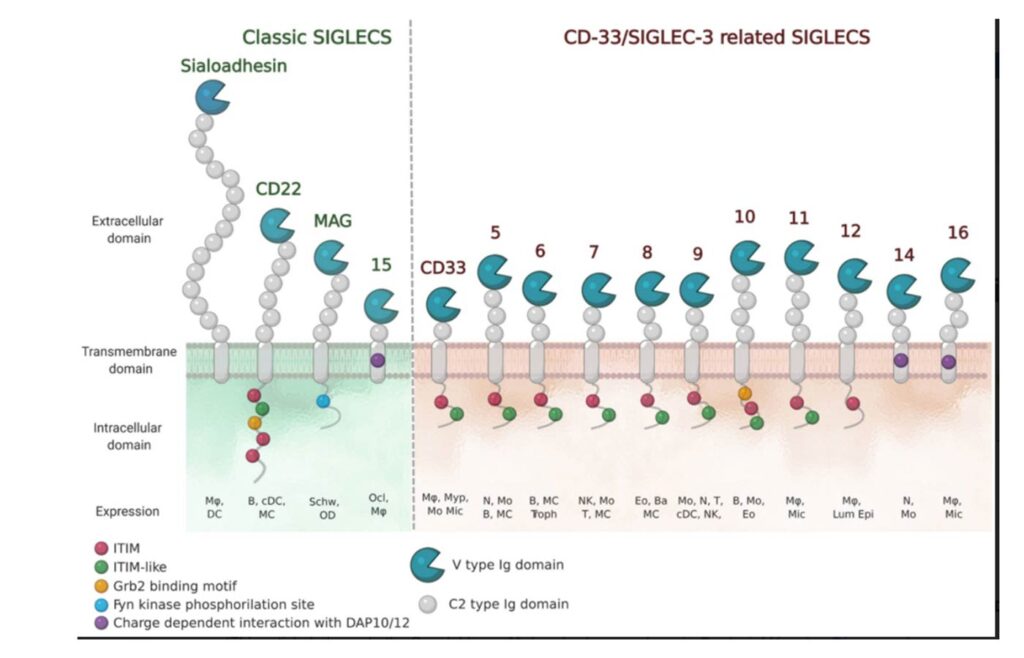
The numbering of Human Siglecs follows the order of their discovery. The representatives murine Siglecs that are not homologous to the human ones are designed by a letter (e.g. Siglec-F). (Varki et al., 2015) Human Siglecs are divided into two groups depending on their homology and evolutionary conservation (Figure 2). CD22-related Siglecs share only 25-30% […]
Sialoadhesin
Sialoadhesin (also known as Sn, Siglec-1 or CD169) was one of the first proteins of the Siglec family to be characterized.(Crocker et al., 1994; Kelm et al., 1994) It shows a high level of conservation between mouse and human, especially in the extracellular region. It is the only Siglec that presents 17 Ig-like domains outside […]
CD22 (Siglec-2)
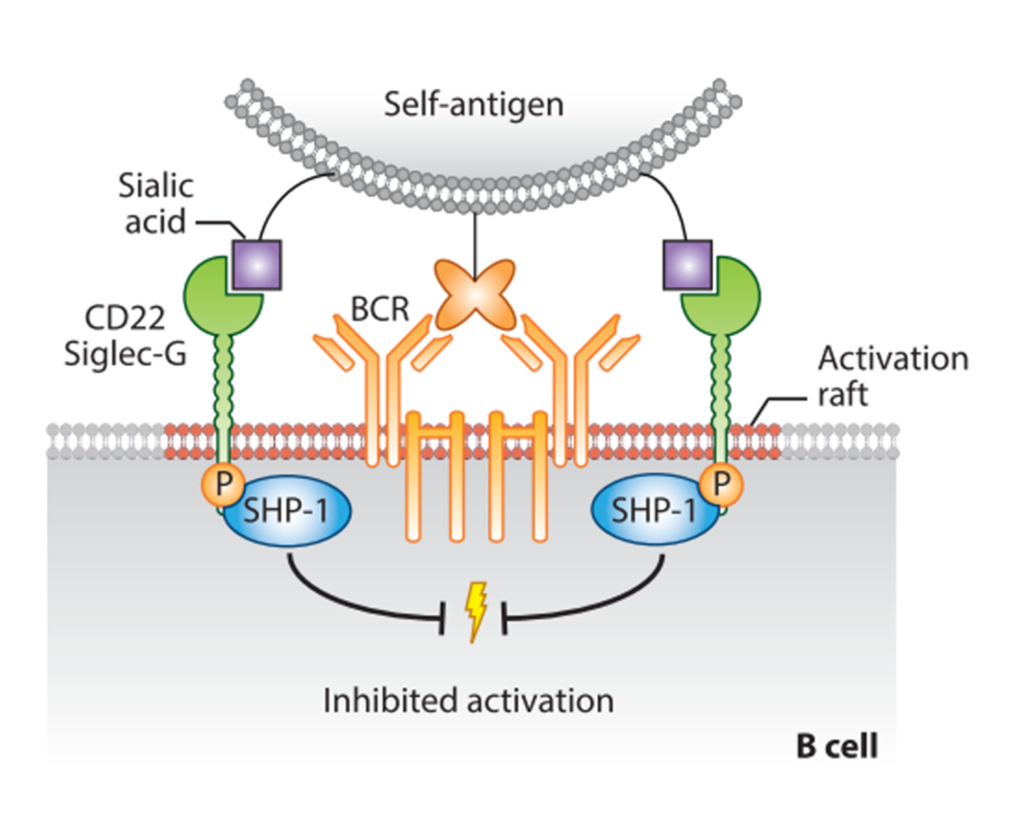
CD22 (Siglec-2) is one of the best described Siglecs. It is well conserved across mammals, and it is highly expressed on B cells, where it regulates their function. There are seven Ig-like domains in the extracellular region, where the V-set domain contains the sialic acid-binding site, as in every Siglecs. The cytoplasmatic tail contains five […]
CD33 (Siglec-3)
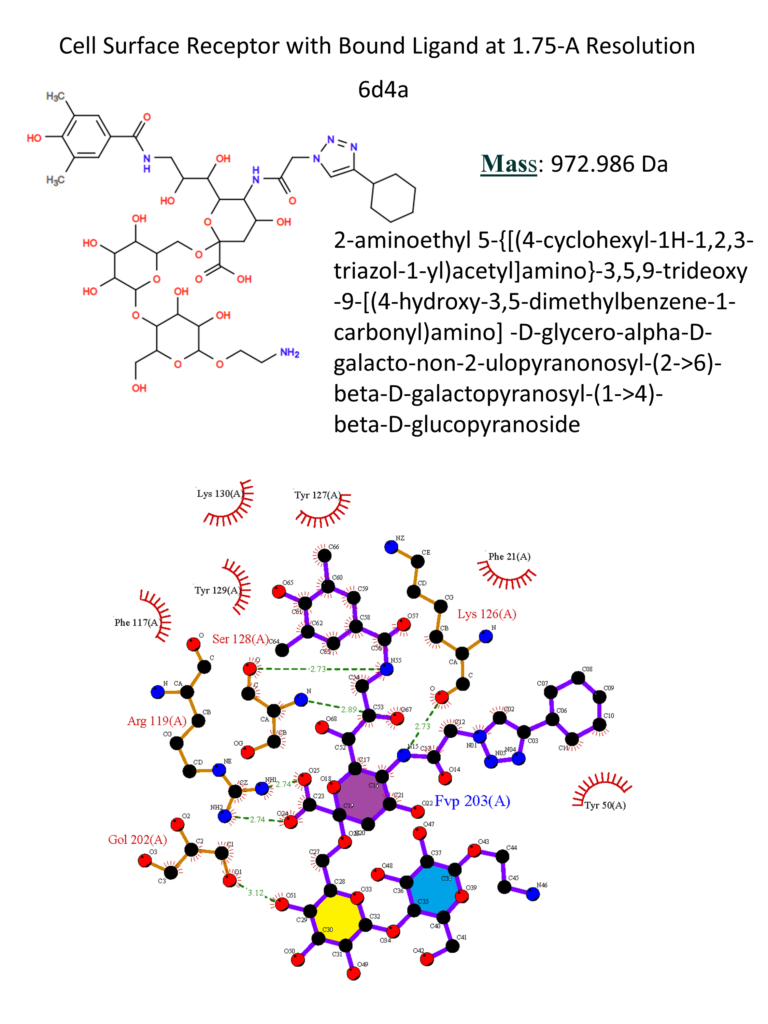
CD33 (Siglec-3) is one of the first Siglecs discovered, and it is the shortest member of the Siglecs family. The extracellular portion presents a V-set domain, which mediates the interaction with the sialic acid, and one C-2 Ig domain: because of its short length, it is more prone to bind ligands on the same cell […]
MAG (Siglec-4)
MAG (Siglec-4) is the most conserved among Siglecs and, unlike the rest, does not have any involvement in the immune system. Indeed, it occurs on oligodendrocytes and Schwann cells in the central and peripheral nervous systems. (Lopez, 2014) It is a highly glycosylated protein, presenting one V-set domain and four C2-set domains in the extracellular […]
Siglec-5 and Siglec-14
Siglec-5 and Siglec-14 are two members of the Siglecs family pair receptors. They occur on the same cells, playing different roles: while Siglec-5 is an inhibitory receptor, Siglec-14 is an activator. Similar to the other Siglecs, the inhibitory activity is associated with the ITIM motif in the intracellular region. Siglec-14 is paired with the activating […]
Siglec-6
Siglec-6 is a member of the Siglecs family expressed on mast cells, B cells, and, notably, on placental trophoblasts, where it may slow down the human birth process. The expression on these last cells is human-specific. (Brinkman-Van Der linden et al., 2007) Siglec-6 contains a V-set domain, two C2-set domains, and one ITIM domain in […]
Siglec-7
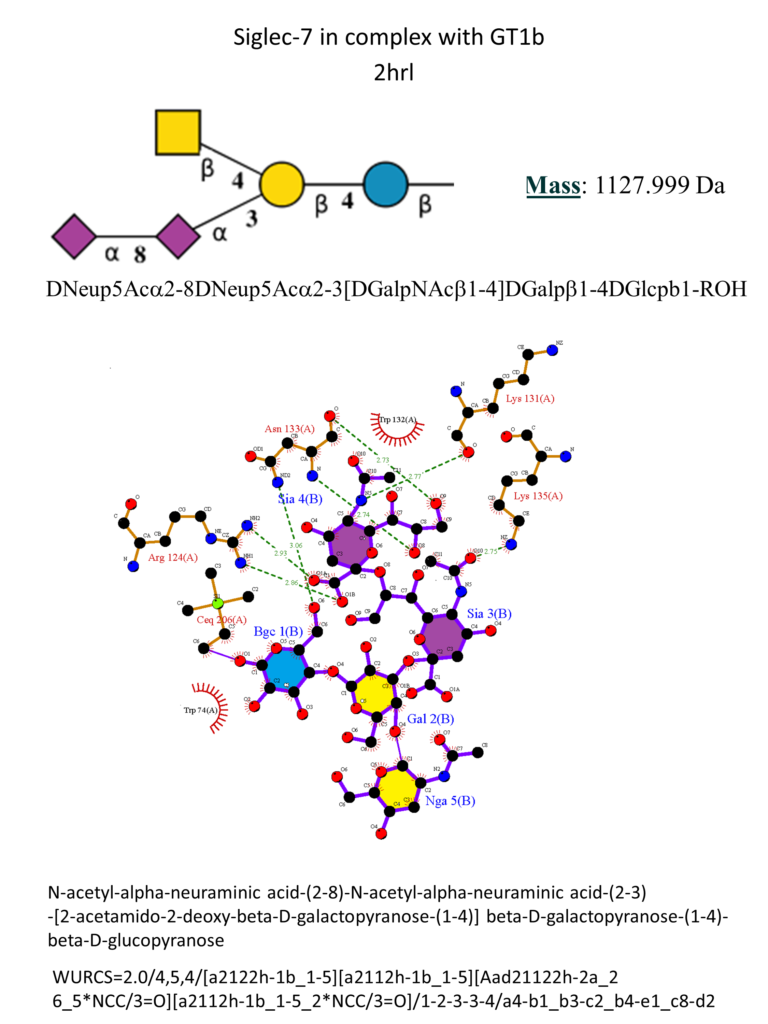
Siglec-7 (also known as p75/AIRM1 or CD328) is a human CD33-related Siglec, described as an inhibitory receptor primarily expressed on natural killer (NK) cells and present on monocytes, dendritic cells, granulocytes and some CD8+ T cells.(Gieseke et al., 2012; Maggi & Moretta, 2020; Shao et al., 2016) It shares high sequence homology with Siglec-9. (Fraschilla […]
Siglec-8
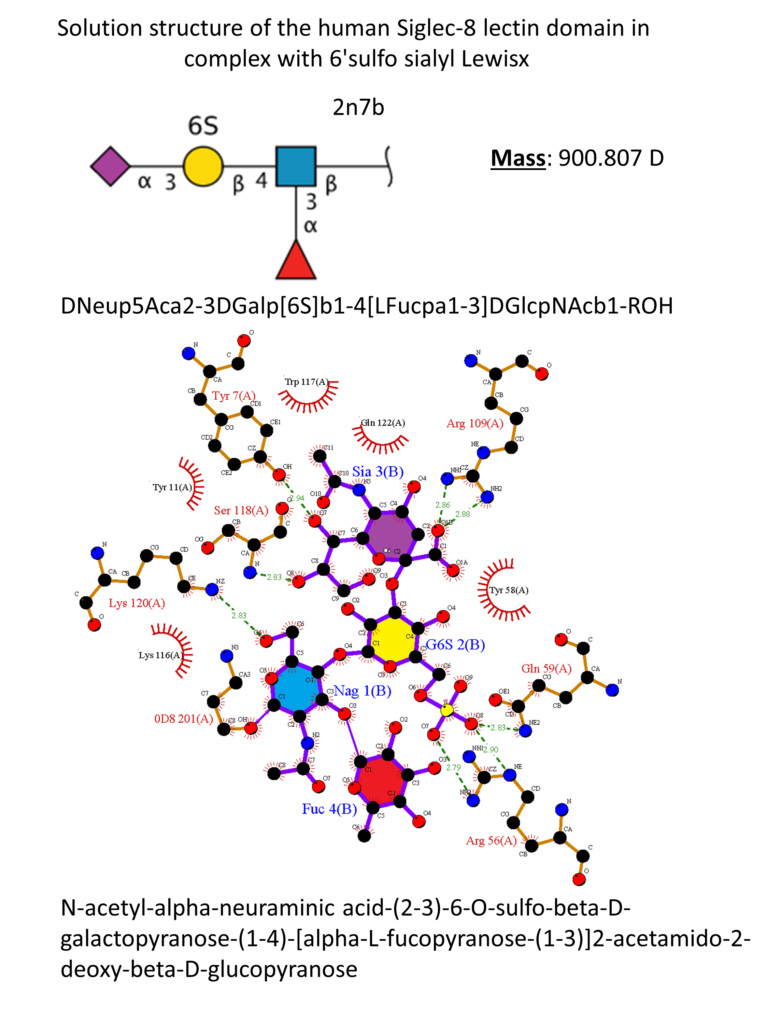
Siglec-8 was discovered around 2000 by two groups independently and identified as a Siglec protein exclusively expressed on eosinophils and mast cells and weakly on basophils.(Floyd et al., 2000; Kikly et al., 2000) Siglec-8 consists of a V-set domain, two C-2 set domains, one ITIM and one ITIM-like domain in the intracellular region. It is […]
Siglec-9
Siglec-9 occurs in human blood on neutrophils, monocytes, plasma cells and a subset of NK cells. It is an inhibitory protein, thus presenting in the cytoplasmatic tail the two putative ITIM domains, while on the extracellular portion, the standard V-set domain and two C-2 set domains. (Zhang et al., 2000) Considering its expression and the […]
Siglec-10
Siglec-10 is an inhibitory receptor expressed on B cells, dendritic cells and leukocytes. Its structure comprises five extracellular domains, two tyrosine-based motifs in its cytoplasmic tail and one Grb2 binding motif close to the membrane.(Duan & Paulson, 2020; Li et al., 2001) Recent studies showed how tumor-associated macrophages present high levels of Siglec-10, which interacts […]
Siglec-11 and Siglec-16
As with Siglec-5 and Siglec-14, Siglec-11 and Siglec-16 form a pair of receptors with opposing functions: Siglec-11 is an inhibitory receptor while Siglec-16 is an activating one. They are both expressed in tissue macrophages and microglia. The expression of Siglec-11 on brain microglia is specific to humans. (Angata et al., 2002; Cao et al., 2008b) […]
Siglec-15
Siglec-15 is a high evolutionary conserved Siglec, characterized in 2007. (Angata et al., 2007) It exclusively occurs on myeloid cells and osteoclasts. Structurally, it only presents one V-set domain and one C2-set domain, while in the intracellular region is coupled, through the positively charged Lys274, with the activating co-receptors DAP12 and DAP10. Unlike the other […]
Conclusions
The idea behind this chapter was to give a general and comprehensive overview of the most recent discoveries about Siglecs, a family of proteins that emerged in the last years as promising pharmacological targets for many diseases. The sialic acid, recognized by Siglecs, is the most abundant sugar of the glycocalyx. It is involved in […]
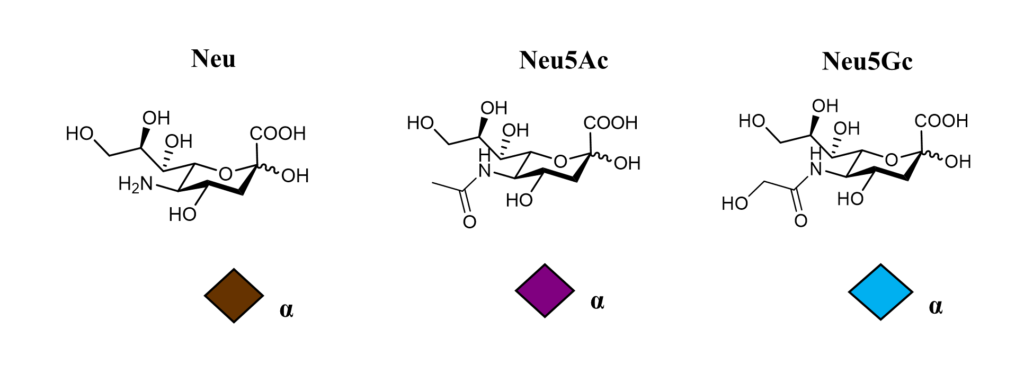
Annex

N-Acetyl Neuraminic Acid α-D N-Glycolyl Neuraminic Acid α-D Neuraminic Acid α-D
The Authors

Benedetta Girardi Benedetta Girardi graduated in Pharmaceutical Chemistry and Technology at the University of Trieste (Italy) in October 2017. She carried out her master thesis with an Erasmus fellowship at the Medicinal Chemistry Laboratory of the University of Complutense in Madrid during her studies. She worked on developing inhibitors of the bacterial protein FtsZ under […]
Abstract
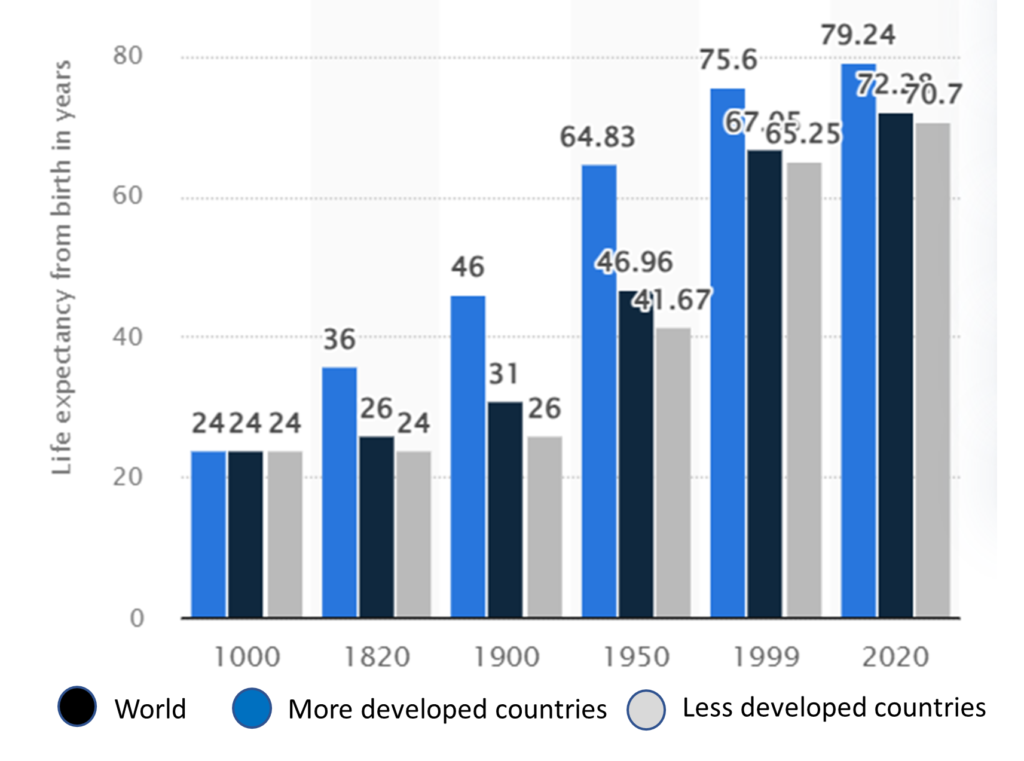
Throughout most of history, average life expectancy from birth was fairly consistent around the world, at about 24 years. This was largely due to high infant and child mortality rates. Those who survived into adulthood could expect to live into their 50s or 60s, but pandemics, food insecurity, and conflict caused periodic spikes in mortality […]
Domestication of Starch
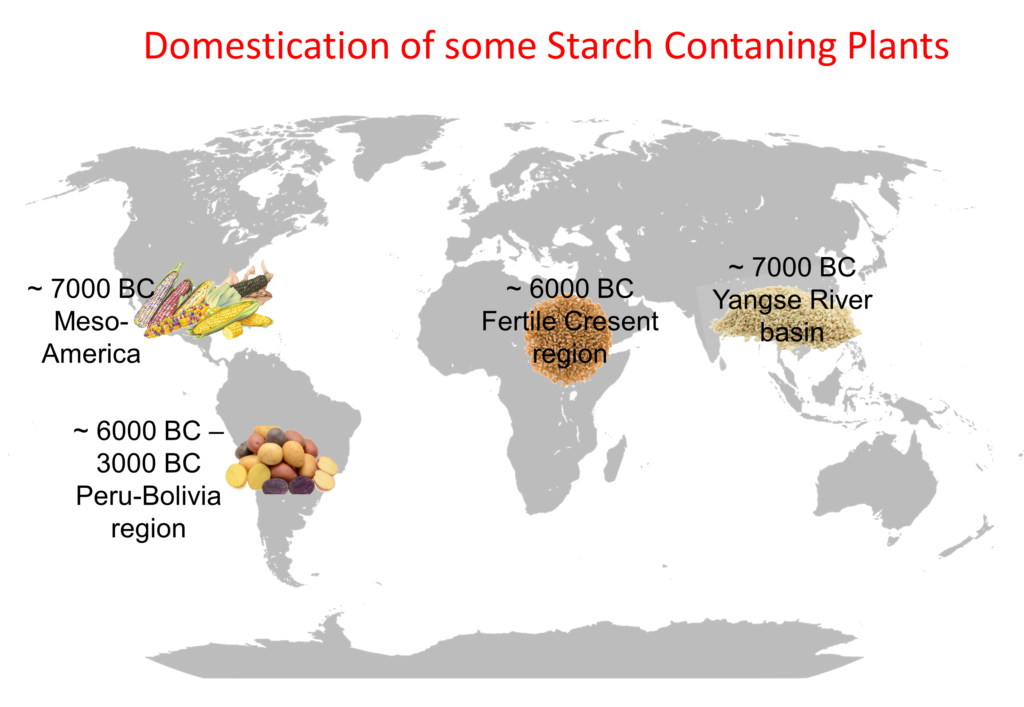
In the past two centuries, we have made monumental advances in nutrition, allowing us to virtually eliminate famine in the developed world. These 200 years or so are just a minute episode in the long co-evolution that humans and plants underwent. Within such a relationship, the most significant advanced development occurred about 15,000 years ago […]
Human Adaptation
The ability to domesticate and cultivate plants close to where humans lived changed how the environment was occupied. As food became more reliable, permanent settlements were established, and the population grew. Early farmers selected the best plant to grow to improve the yield and secure crop production. They continued this process generation after generation until […]
A Brief History of Potatoes
The history of potatoes throughout their domestication, exportation, implementation and contribution to contemporaneous society illustrates the uniqueness of starch as a food, materials and energy unique renewable biological resource. Today, the potato is the fifth most important crop worldwide, after wheat, corn, rice and sugar cane. Potatoes were first domesticated in the Andean highlands as […]
Non-Food Usages
Although the early history of starch usage is mainly unrecorded, some very early examples exist of its industrial use. In the pre-dynastic period (4000 BC), Egyptians cemented strips of papyrus together with starch adhesive made from wheat. At a later date, Chinese documents were first coated with a high-fluidity starch to provide resistance to ink […]
Unveiling the Starch Structure

Starch has been used over several millennia for many different applications. However, research on understanding this substance spans about three centuries starting with Leeuwenhoek, who observed it microscopically as discrete granules in 1716. About 250 million years ago, the world consisted of a single giant landmass now known as Pangaea. Geological forces broke Pangaea apart, […]
Starch in the Global Bioeconomy
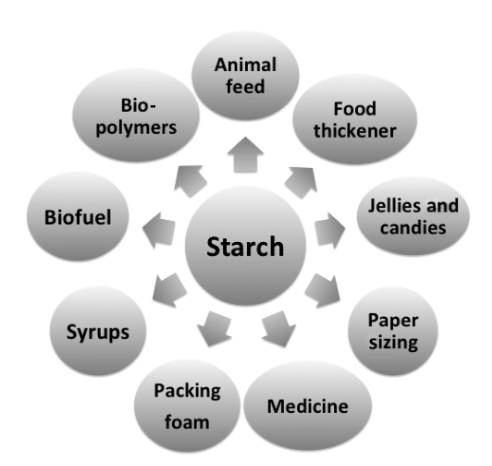
More than ever, starch remains an irreplaceable pillar of our society. It is the main storage polysaccharide in foods of plant origin and the primary energy source in the human diet. Cereals (such as wheat, corn, rice and barley), tubers and roots (such as potatoes and cassava) are the sources of common food starches. Starch […]
References
This article is an upgraded version of the Preface written for the book “Starch in the Bioeconomy”, by J-L. Wertz & B. Goffin” , (2021) Taylor & Francis Group. Schaal B. Plants et al., Our shared history and future. Plants, People, Planet, 2019;1:14–19. https://doi.org/10.1002/ppp3.12 V.K. Vimmwock & J.N. BeMiller. The architecture, nature, and mystery of […]
Movies

The wonderful tales of potatoes (Movie 22minutes) Mais/Corn/Korn (Movie 3m40)
Abstract
This chapter offers a general background for students embarking in glycoscience to grasp an essential component of the field, i.e. the alphabet of the building blocks that constitute the many naturally occurring glycans and complex carbohydrates. There is a need to conform to the recommendations of nomenclatures of carbohydrates whilst the constraints required by the […]
Nomenclature
Monosaccharides are the chemical units from which all members of the major family of natural products, the carbohydrates, are built. They are the individual carbohydrate building blocks, i.e. the monomeric constituents of more complex architectures that will be referred to as glycans, an assembly of sugars either in free forms or attached to another molecule […]
At the Service of Glycobiology
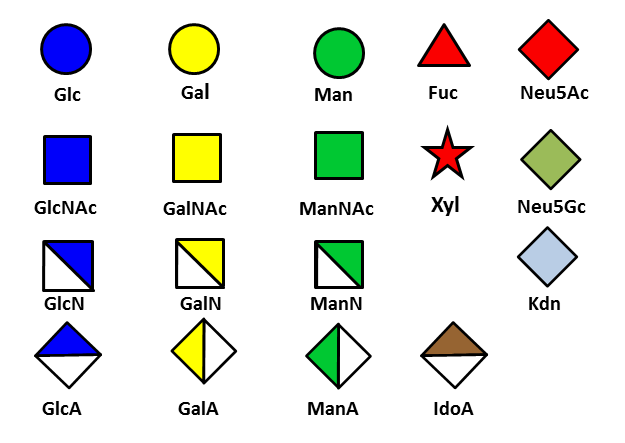
The multiple monosaccharide building blocks can be linked to various regio-chemistries and stereo-chemistries, and the resulting glycan be assembled on protein or lipid scaffolds. The diversity of glycoconjugate structures comprises an “information-rich” system capable of participating in a wide range of biological functions. There comes the question of representing and encoding the many monosaccharides in […]
Beyond the Symbols ….
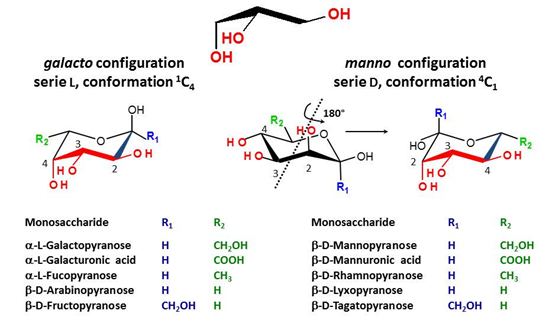
It is fully recognized that no symbol system will ever convey a full appreciation of the three-dimensional structures of glycans, essential information for the understanding of how glycans and proteins interact (A. Varki et al., 2008). This is clearly illustrated by the example given in Fig. 11 which shows the many monosaccharide units that can satisfy […]
150 Monosaccharides
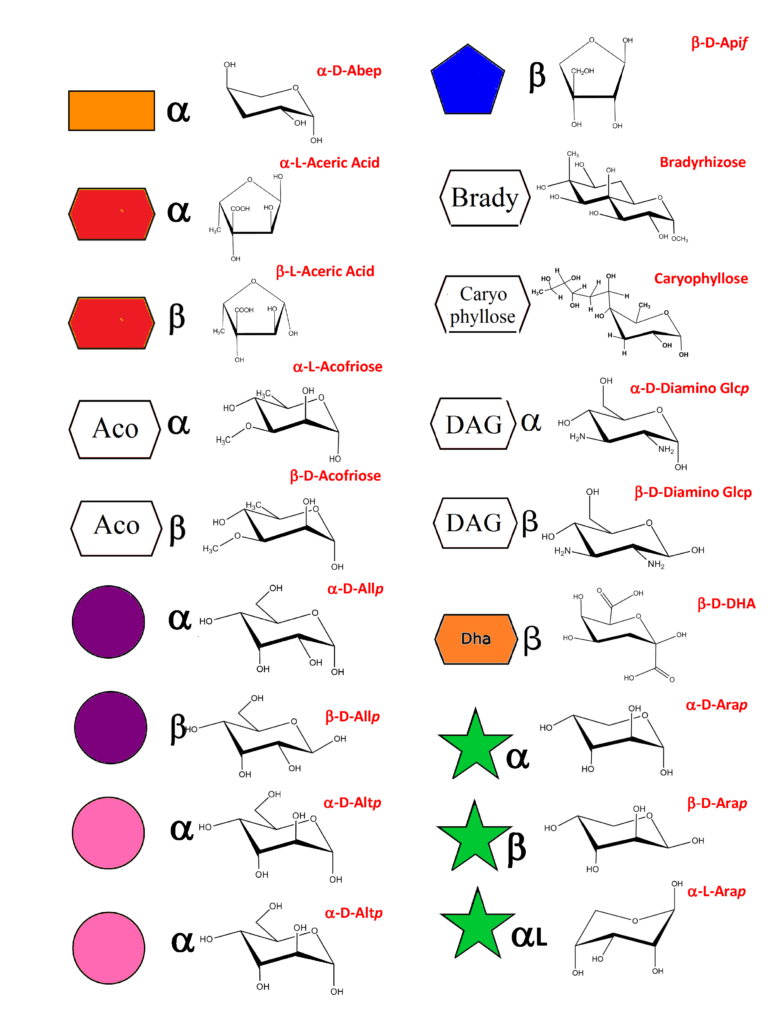
While maintaining the spirit of using the symbolic representation for monosaccharides (and towards glycans) this set of rules provides the necessary extension to the construction of three-dimensional structures, allowing encoding for computational manipulation whilst maintaining IUPAC nomenclature. As a result of an extensive search, more than 150 monosaccharides have been identified as the building blocks […]
References
Berger, O., McBride, R., Razi, N. & Paulson, J. (2008) “Symbol Notation Extension for Pathogen Polysaccharides”, The Scripps Research Institute, Consortium for Functional Glycomics. Bijvoet, J.M., Peerdeman, A.F. van Bommel, A.J. (1951) “Determination of the Absolute Configuration of Optically Active Compounds by Means of X-rays”, Nature, 168, 271-272. Consortium for Functional Glycomics : http://www.functionalglycomics.org Engelsen, S.B., Hansen, […]
How do Lectin Read the “Glyco Code”
Of all the building blocks of life, glycans are the most abundant and diverse family of organic molecules, which informational content potentially exceed that of nucleic acids, proteins and lipids. Their complexity originates from the innumerable regio- and stereo-diverse ways in which simple sugar monomers can assemble to form highly branched polymers. This dense array […]
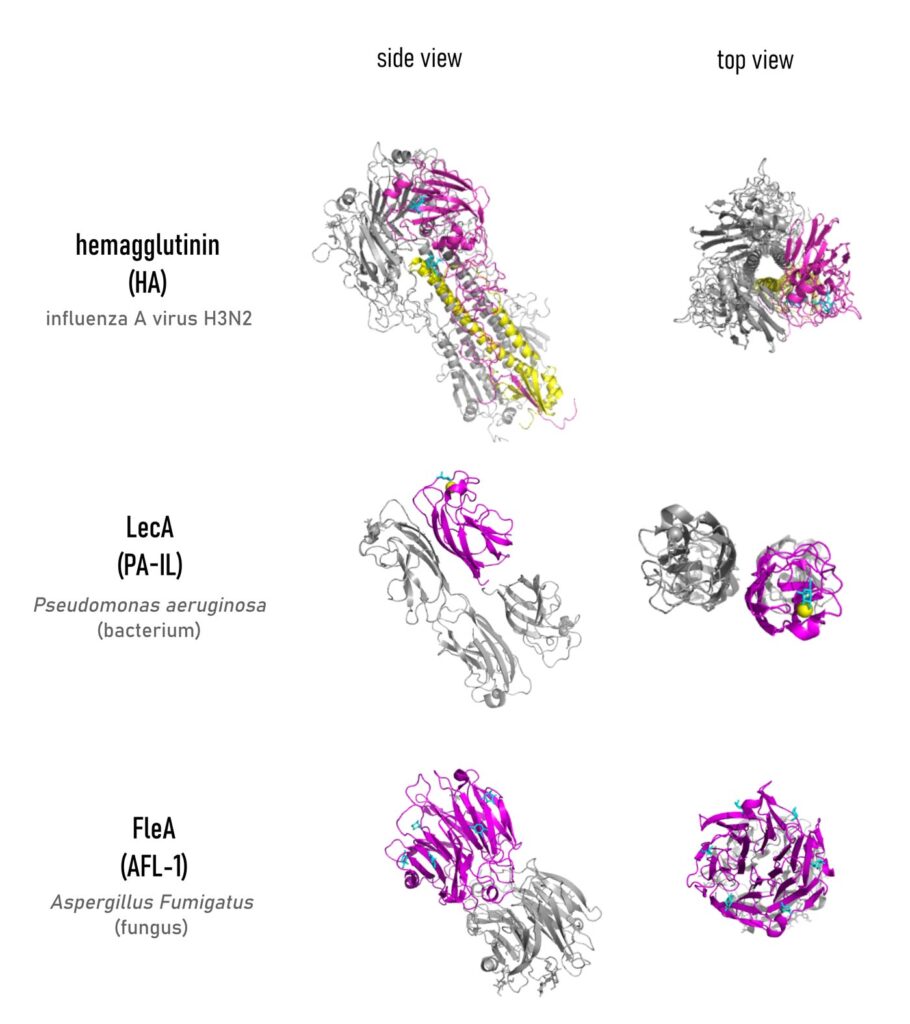
Understanding Multivalency
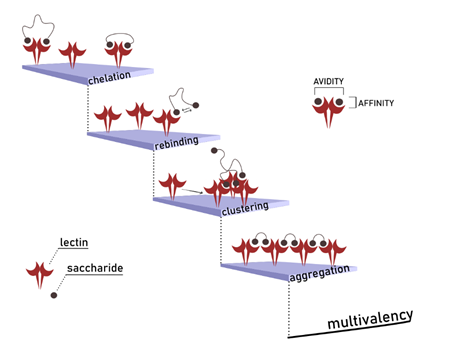
Often possessing more than one binding site, lectins are thus intrinsically multivalent receptors that can interact with multiple copies of a monovalent ligand at once. For instance, a high display of carbohydrate epitopes is represented by the glycocalyx, which is the complex carbohydrate coating that decorates the cellular surface. The glycocalyx is an illustration of […]
Measuring Carbohydrate-Lectin Interactions

When studying the role of lectin in biology or when assessing the capability of new therapeutics in (dis)regulating these functions, the choice of an assay suitable for this purpose is of absolute importance. Along with the correct set up of a carbohydrate-protein binding assay, an appropriate interpretation of the outcome of such experiments is a […]
In solution Assays: Isothermal Titration Calorimetry
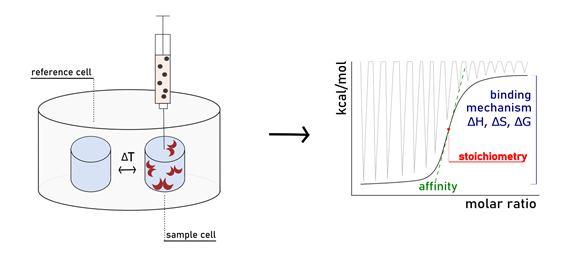
Isothermal Titration Calorimetry (ITC) is considered the method of choice to analyse protein-carbohydrate interactions. It allows the precise determination of the thermodynamic parameters associated with the binding process (ΔG, ΔH, ΔS, Kd) in a single experiment. This technique consists of titrating a protein solution with that of a ligand, or vice versa. Immediately after the […]
In Solution Assays: Fluorescence Polarization
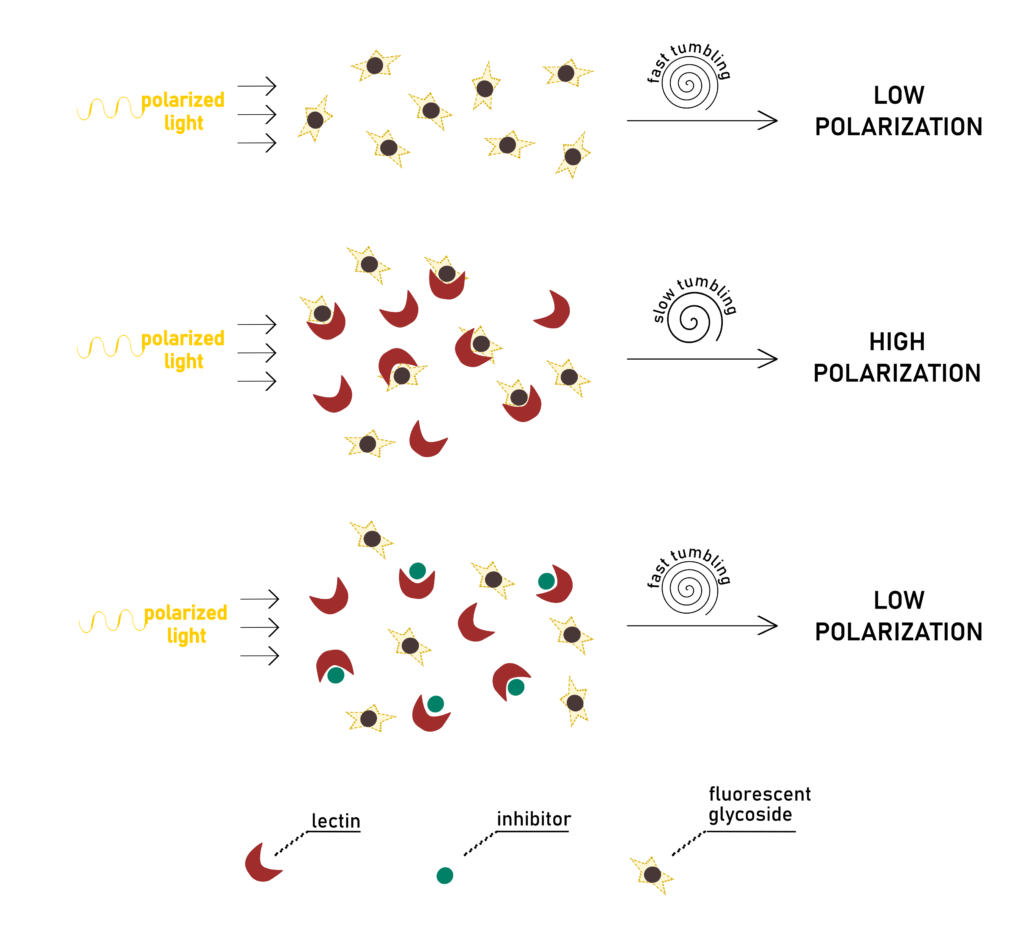
Fluorescence polarization (FP) is a powerful tool in biochemical research. This method has historically been applied to the study of protein-protein and acid nucleic-protein interactions (Moerke, 2009; Checovich et al., 1926). It is also a suitable method for the screening of drug candidates in the inhibition of enzymes or receptors, such as GPCRs ()Lea & […]
In Solution Assays: Hemagglutination Inhibition
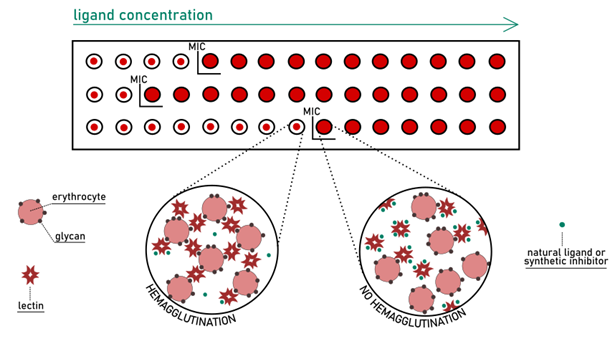
In the early history of lectin research, the most common method to identify lectin activity was to check the ability to agglutinate erythrocytes. In the past, lectins were referred to as “agglutinins” because of their ability to cause cell agglutination through binding the sugar residues present on the surface. Nowadays, it is known that such […]
On Surface Assays: Enzyme-Linked Lectin
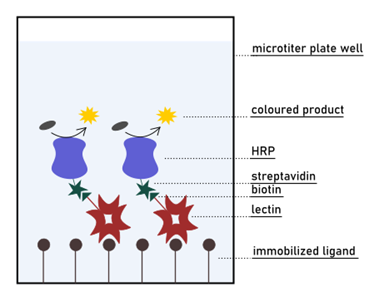
The enzyme-linked lectin assay (ELLA) was introduced in 1983 by McCoy et al. (. McCoy et al., 1983 ) as a variation of the more famous enzyme-linked immunosorbent assay (ELISA). The methodology was adapted to the evaluation of binding properties of lectin ligands, as a valid alternative to the widely used HIA. ELLA is a quantitative technique […]
On Surface Assays: Surface Plasmon Resonance
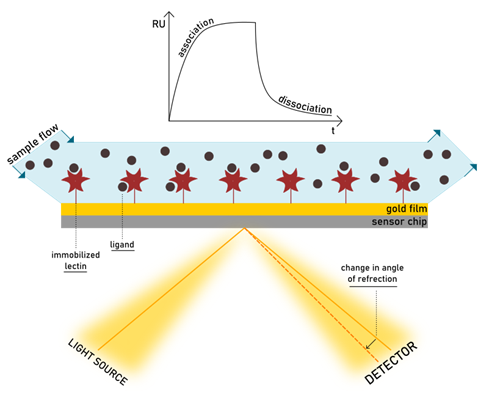
The phenomenon of surface plasmon resonance (SPR) was observed for the first time by Wood more than a century ago (Wood, 1902). Since then, the number of publications on the topic increased exponentially (Rich et al., 2011), until reaching several thousand today. Only 90 years after the discovery of the theory of SPR, the first […]
Conclusions and Outlook
The relevance of carbohydrate-lectin interactions in controlling many physiological processes is key to various disciplines. However, the understanding of the molecular basis regulating such events is still limited. The main take-home message that we want to convey with this e-chapter is that the choice of a correct assay is fundamental for the study of binding […]
Bibliography
Andre, S.; Pieters, R. J.; Vrasidas, I.; Kaltner, H.; Kuwabara, I.; Liu, F. T.; Liskamp, R. M. J.; Gabius, H. J. Wedgelike Glycodendrimers as Inhibitors of Binding of Mammalian Galectins to Glycoproteins, Lactose Maxiclusters, and Cell Surface Glycoconjugates. ChemBioChem 2001, 2 (11), 822–830. https://doi.org/10.1002/1439-7633(20011105)2:11<822::AID-CBIC822>3.0.CO;2-W. Audfray, A.; Varrot, A.; Imberty, A. Bacteria Love Our Sugars: Interaction […]
Abstract
Multi-drug resistant (MDR) pathogens such as Burkholderia cenocepacia have become a hazard in the context of healthcare-associated infections, especially for patients admitted with compromising or aggravating conditions. Like other opportunistic Gram-negative bacteria, this pathogen establishes virulence and biofilms through lectin-mediated adhesion. Glycans and glycomimetics have become devices of choice to antagonize or disrupt such interactions. We provide an […]
The problem of antimicrobial resistance (AMR)
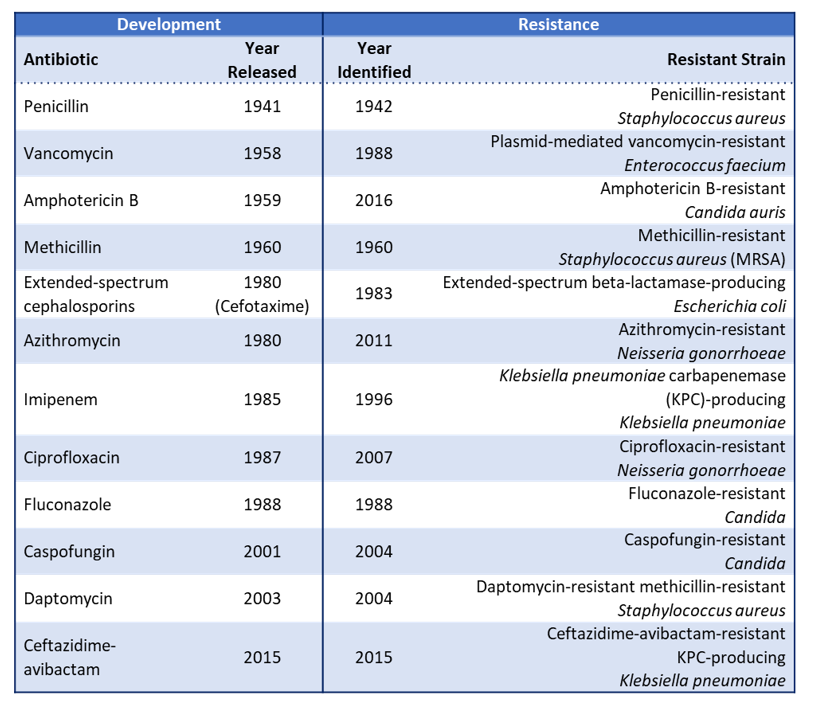
The problem of antimicrobial resistance (AMR) Many barriers have been met and overcome across the history of the human race and its progress. One of such, and particularly significant, is humans’ fight against pathogenic microorganisms. Very relevant to current times, pathogenic viruses can rise to become global threats, but so can bacteria. Be it the […]
Anti-adhesion therapy (AAT), a possible solution
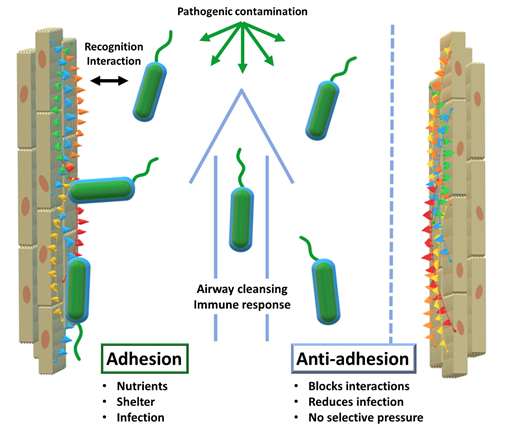
In order to act efficiently, infective pathogens need to interact with their environment. First and foremost, a virus or a bacterium needs to recognize the cells of its host to start the infective process. At this point, it becomes necessary for the pathogen to remain in close vicinity to its host cells. In this vicinity, […]
The role of lectins and carbohydrates in infection
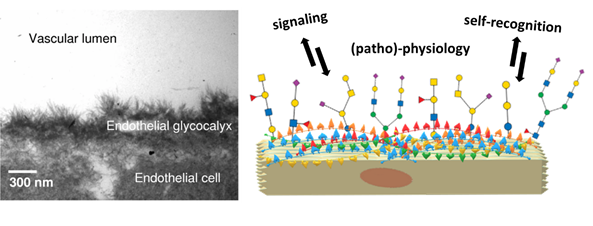
Human epithelial cells are found at the forefront of human anatomy separate the body and its cavities from the exterior environment. Their glycocalyx nanolayer comprises glycoconjugates: glycoproteins and glycolipids, which present their carbohydrate portion to the extracellular environment. The role of the glycocalyx and its actors is to sense and communicate with their environment in […]
Glycans and Glycomimetics as therapeutic agents
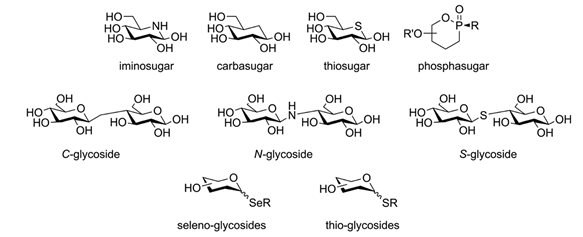
On paper, the concept of using naturally occurring carbohydrates in AAT can be considered a revamping of the successful story involving breastfeeding and HMOs. Unsurprisingly, it was implemented as early as the late 70s and was successful to an extent: in vitro and in vivo experiments repeatedly prevented infections in models featuring common pathogens such […]
Opportunistic lung pathogens: Burkholderia cenocepacia and company

As previously mentioned, multidrug-resistant (MDR) pathogens are a constant threat to hospitalized patients, especially those with risk factors such as cancer, diabetes, immunodeficiency, etc. Indeed, opportunistic pathogens take advantage of their weakened organisms for infecting and spreading among patients, leading to outbreaks of hospital-acquired infections (HAIs). Among MDR opportunistic pathogens, lung pathogens are especially notorious: […]
Lectins of B. cenocepacia: the BC2L family
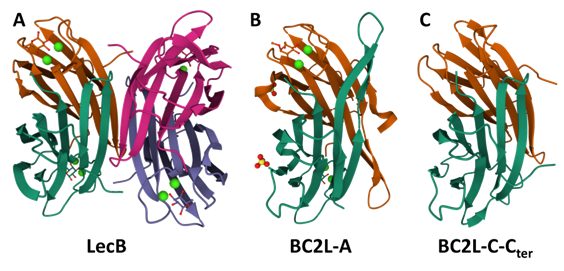
As mentioned earlier, lectins are key actors in cell adhesion leading to infection and have proven to be interesting anti-adhesion and combination therapy targets. A prime example of these notions is how inhibiting the soluble lectins of P. aeruginosa with drug-like glycomimetics has led to biofilm disruption and enhanced susceptibility to antibiotics. (Sommer et al., […]
The superlectin BC2L-C: state of the art
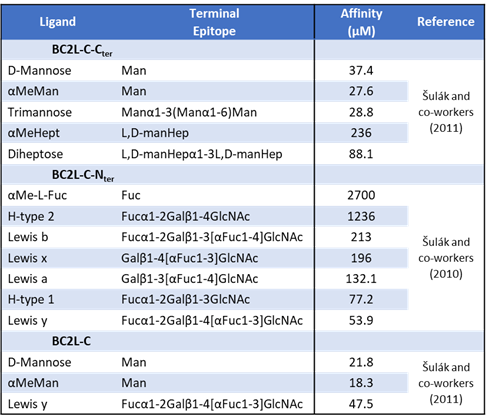
Like BC2L-A, the protein BC2L-C is a “Lec-B” like lectin: its C-terminal domain is 116 residues long and shares 43% identity with LecB. (Sulak et al., 2011) Much like BC2L-A, this domain assembles itself as a homodimeric lectin, featuring two calcium-dependent binding sites. Continuing the similarities, the specificity loop in the binding site bears alanine […]
References
Conclusion
In conclusion, two undeniable truths must be considered: antimicrobial resistance is becoming a critical problem for modern medicine, and the use of glycomimetics to fight against drug-resistant infections holds great potential. Indeed, the roles played by lectins and carbohydrates during the infective process makes them excellent targets for disrupting and preventing infections. Moreover, scientific and […]
The Author

Rafael Bermeo Born and raised in Ecuador, Rafael Bermeo moved to France in 2011 to attend the Lyon Superior School of Chemistry, Physics and Electronics (CPE Lyon), where he obtained his Bachelor, then his Chemical Engineering Diploma in 2018. During his diploma, Rafael spent one year as an exchange graduate student at Oregon State University […]
Introduction
The immune system has evolved to an amazing defense mechanism against the invasion of a broad variety of pathogens and parasites. It is possessed by the majority of the living organisms including such simple beings as unicellular bacteria or invertebrates, where it exists in a rudimental form (He, J & Deem, M. W. 2010, Rowley, A. […]
The non-specific and specific components of innate immunity
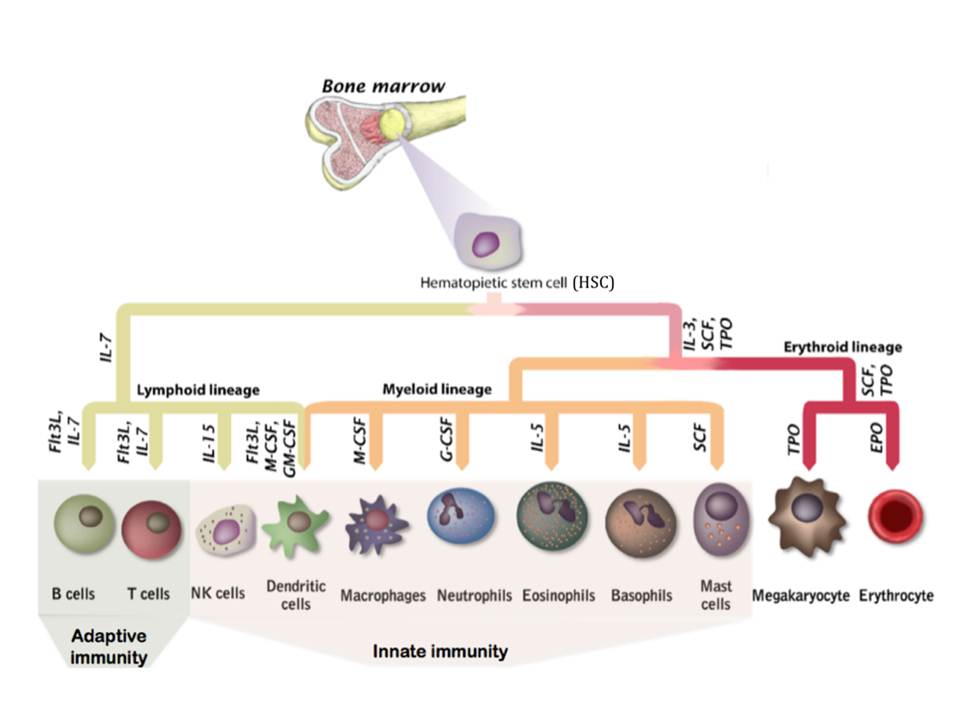
Anatomic and chemical-physiological barriers The skin is composed of two layers, epidermis and dermis. The epidermis consists of several layers of tightly packed cells, where the outer layer contains dead cells and waterproof protein keratin, and most of the pathogens are not capable to cross such a barrier. The underlying dermis contains sebaceous glands that […]
Pathogen recognition by innate immunity
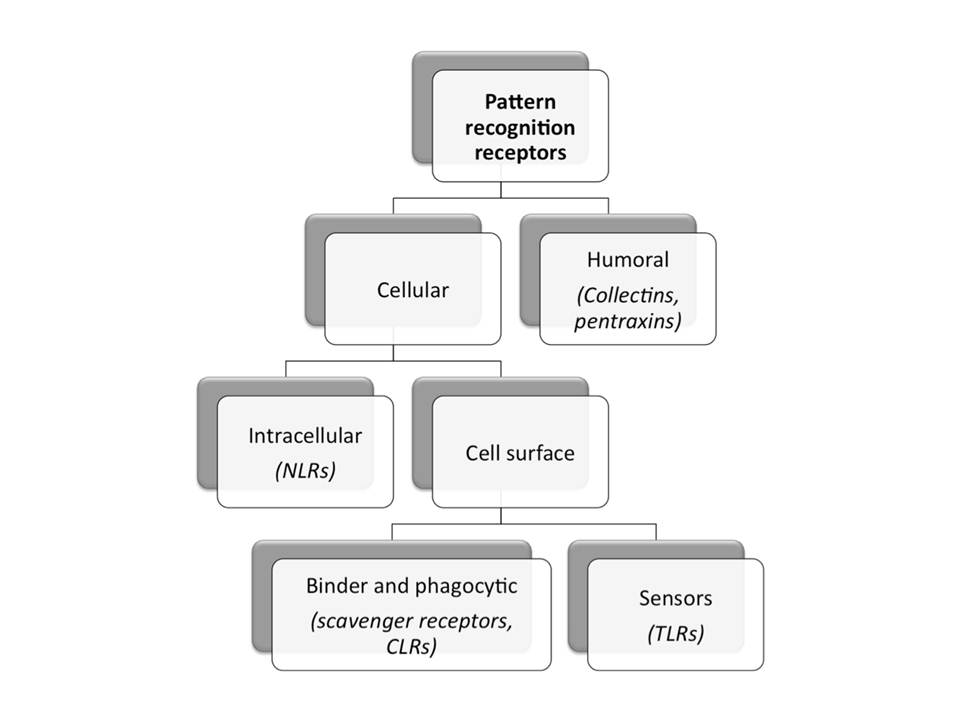
The innate immune system functions using at least two recognition strategies: it is capable to distinguish microbial non-self from missing self. These recognition events occur thanks to a range of receptors that all cells of the innate immune system are equipped with, and through which various signals can be triggered by cytokines, conserved components of […]
Dendritic cells and adaptive immunity initiation
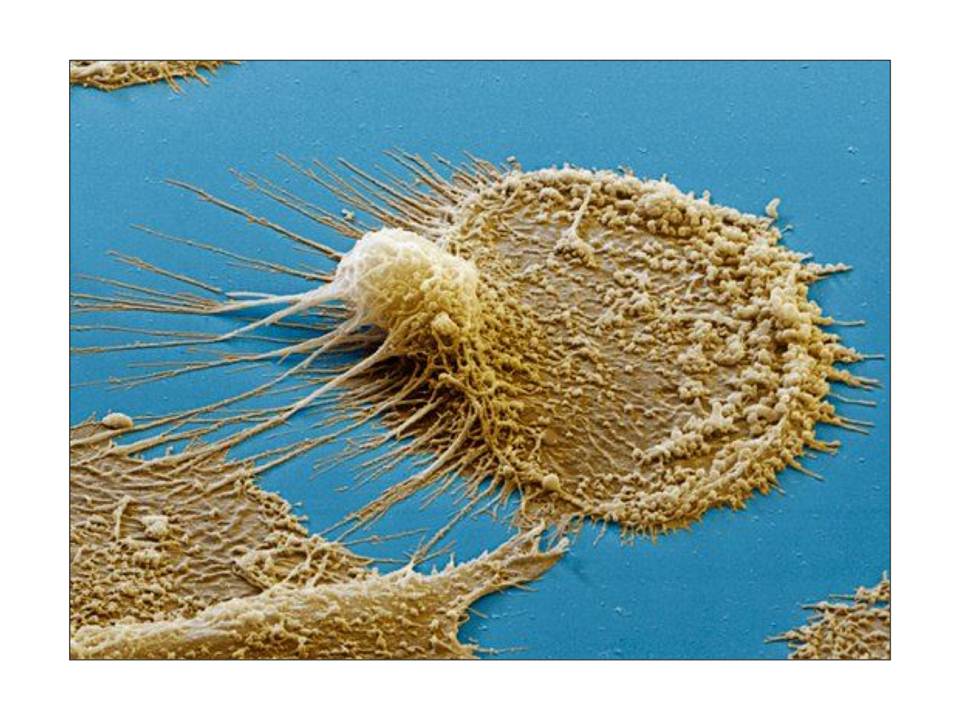
Dendritic cells comprise a diverse group of professional antigen presenting cells (APCs), which share a particular morphology, i.e. long surface membrane extensions called dendrites (fig. 8), and have a common feature of being potent stimulators of T cells to induce the adaptive immunity. The diversity of DCs is a result of a combination of several […]
References
Bashirova, A. A, Geijtenbeek, T. B, van Duijnhoven, G. C, van Vliet, S. J, Eilering, J. B,Martin, M. P, Wu, L, Martin, T. D, Viebig, N, Knolle, P. A, KewalRamani, V. N, van Kooyk,Y, & Carrington, M. (2001) A dendritic cell-specific intercellular adhesion molecule 3-grabbingnonintegrin (dc-sign)-related protein is highly expressed on human liver sinusoidal endothelial cells and […]
Authors
Ieva Sutkeviciute After graduating as a Master of Science (Biochemistry, Molecular Biology, and Biopharmacy) from the Vilnius Gediminas Technical University, Ieva spent three years as an analyst working within the company : Sicor Biotech – TEVA pharmaceuticals, in Vilnius. There she carried on the evaluation of biopharmaceutical interactions with their receptors using Surface Plasmon Resonance. In […]
Introduction
Why are glycans so important ? Glycans, also known as carbohydrates, saccharides or sugars, cover all living cells from humans to virus going through plants or bacteria, among others. These features make glycans the first contact point in any host-guest interaction, spanning from symbiotic to pathogenic relationships. In humans and animals, they are found on cells […]
Microbial Glycoconjugate Structures
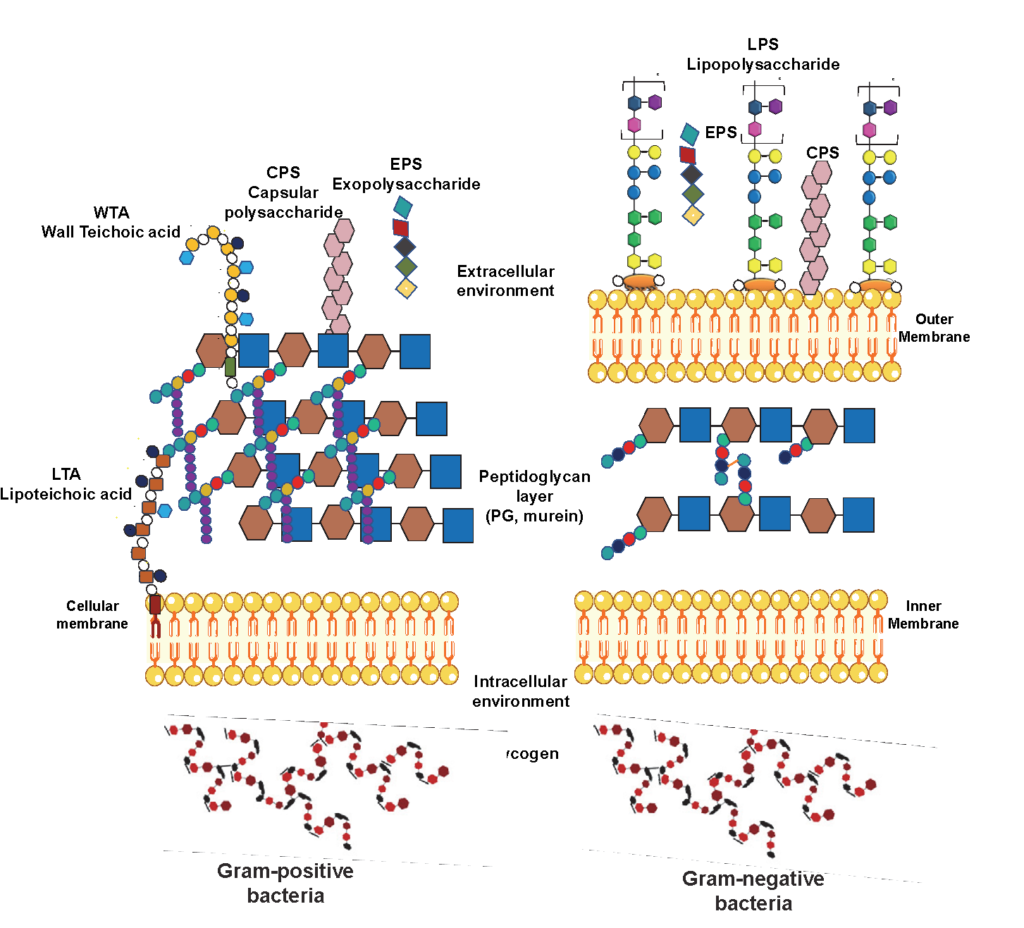
Bacterial polysaccharides can be classified into intracellular or extracellular polymeric substances (Figure 2). On the one hand, there are the intracellular polysaccharides (IPS), the cells’ energy source. On the other hand, the extracellular polymers are subdivided into two main polysaccharide groups: cell wall constituents, such as lipopolysaccharides (LPS), teichoic acids, and peptidoglycans (PG), and polysaccharides […]
Computational Tools for the Structural Elucidation
Determining the glycan structure, presents many challenges, mainly related to glycan’s complexity and diversity, together with the non-templated nature of glycan biosynthesis. Simple sugar monosaccharides can be assembled through linkages at many different sites, forming complex polymers which can adopt a wide variety of shapes. (46) A major difficulty occurs when dealing with bacterial glycans […]
Carbohydrate Databases
First of all, the accumulation and processing of published carbohydrate-related information resulted in several databases, including glycan structures and properties, taxonomic and bibliographic annotations, and experimental, analytical data. The main curated and freely available carbohydrate databases are listed below. GlyTouCan: It is a repository part of the GlyCosmos web portal. Glycan structures can be freely […]
Visualising and Building Glycans
Many different publicly accessible glycoinformatics tools allow the building and visualisation of glycans structures. A thorough description of these instruments is provided in the review of Lal et al., (52). Nevertheless, a list of the main user-friendly programs, software, and servers used for sketching glycans is reported below. SugarSketcher: JavaScript interface to draw online glycan […]
Bioinformatics & Determination of Glycan Structures
Above all, bioinformatics tools are useful for achieving the hard goal of elucidating complex glycans structure, making it easier to interpret the experimental data. As mentioned above, these molecules’ structural analysis implies a complex operation of bench organic and analytical organic chemistry. Only once extracted and purified the polysaccharide material it is possible to proceed […]
Mass Spectrometry Tools
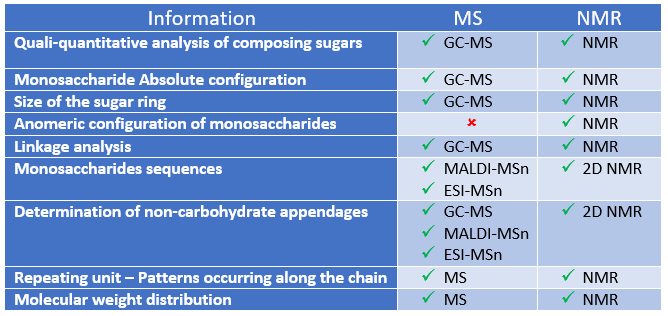
Mass Spectrometry is one of the main analytical techniques currently used in the structural analysis of glycans. In particular, MALDI and ESI MS methods are well suited to detect various glycoconjugates’ glycosylation profiles. It is worth considering that tandem mass spectrometry, often combined with liquid chromatography and applied on complex mixtures, provides a large amount […]
NMR Spectroscopy Tools
The combination of mass spectrometry and NMR spectroscopy represents a powerful tool for the full characterisation of glycans. NMR spectroscopy is indeed a non-destructive method that provides key information useful not only in the elucidation of the chemical structure but also in the conformational analysis of complex glycans, including, for example, the number of the […]
Conformational Analyis of Complex Glycans
The elucidation of complex glycans conformation in solution is one of the main intriguing aspects of bacterial oligo- and polysaccharides’ full characterisation. The function and the biological role of glycans are indeed strictly related not only to their structure but, above all, to their conformation. Even single furanoid or pyranoid monosaccharides can adopt possible shapes; […]
Glycan 3D modeling & 3D Structures Displays
Glycam: Online web that provides tools for 3D modelling carbohydrate-containing structures. It includes libraries and prediction tools together with useful drawing, scripting and building tools. (http://glycam.org/tools/molecular-dynamics/oligosaccharide-builder/build-glycan?id=1) GLYCAM-web (Carbohydrate Builder): Online tool for carbohydrate building through three different options: manual building, using templates or in text format for structures that do not exist in the library. […]
Conclusions
In conclusion, all the bioinformatic tools described here are pivotal support to the interpretation of experimental data and represent a key point for the description of the chemical structure and the conformation of complex glycans. Advances in analytical methods, including Mass Spectrometry- and NMR spectroscopy-based techniques, increased the qualitative and quantitative data of glycans collected in […]
References
1. Aspinall, G. O., The Polysaccharides. Vol. 2 Academic Press : Orlando ; London, 1983 2. Turnbull, J. E. ; Field, R. A., Emerging glycomics technologies. Nat Chem Biol 2007, 3 (2), 74-7. 3. Aoki-Kinoshita, K., A Practical Guide to Using Glycomics Databases. 2017. 4. Barcelos, M. C. S. ; Vespermann, K. A. C. ; Pelissari, F. M. ; Molina, G., Current […]
The Authors
Ferran Nieto Fabregat Ferran graduated in chemistry with a biological mention in 2018 at Universitat Autònoma de Barcelona (Spain), basing his final degree thesis in organic synthesis with carbohydrates. In 2019, after finishing the Master degree, he moved to Naples, where he started a PhD under the Marie Skłodowska-Curie actions fellowship in the Universittà degli […]
Abstract Introduction

https://glycopedia.eu/wp-content/uploads/2019/09/from_chitin_to_chitosan-2-c08.pdf Henri Braconnot, who was the director of the Botanical Gardens at the Academy of Sciences in Nancy, France, discovered chitin in 1811 after the report of a “material particularly resistant to usual chemicals” by A. Hachett, an English scientist in 1799. The substance, named “fungine,” was extracted from mushrooms that would not dissolve in […]
Occurrence of Chitin

Chitin (β-1,4-linked 2-acetamido-2-deoxy-D-glucose) is found throughout the biosphere. Its estimated production is 1010 to 1012 tons per year. Chitin is the second most abundant polymer after cellulose. Sea animals, insects, and micro-organisms are the sources of chitin. In more than 90% of all animal species and insects, chitin-based composites are the major constituents of the […]
Chitin Metabolism
The coordination of chitin synthesis and its degradation re-quires strict control of the participating enzymes for subsequent growth and development in insects and fungi. As for insects, growth and morphology depend on the capability to remodel, in time and space, chitin-containing structures. Insects, repeatedly produce chitin synthases and chitinolytic enzymes in different tissues.Merzendorfer & Zimoch, […]
Extraction of Chitin & Preparation of Chitosan
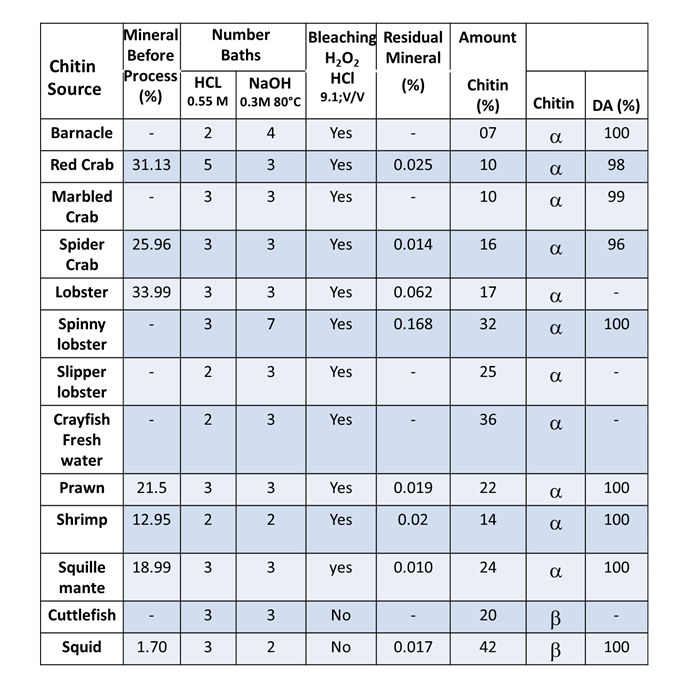
4.1. Extraction of Chitin The primary commercial sources of chitin are crab and shrimp shells. The isolation of chitin begins with the selection of shells, which has an important bearing on the subsequent quality and property of the final isolated material. For example, for lobsters and crabs, shells of the same size and species are […]
Nomenclature
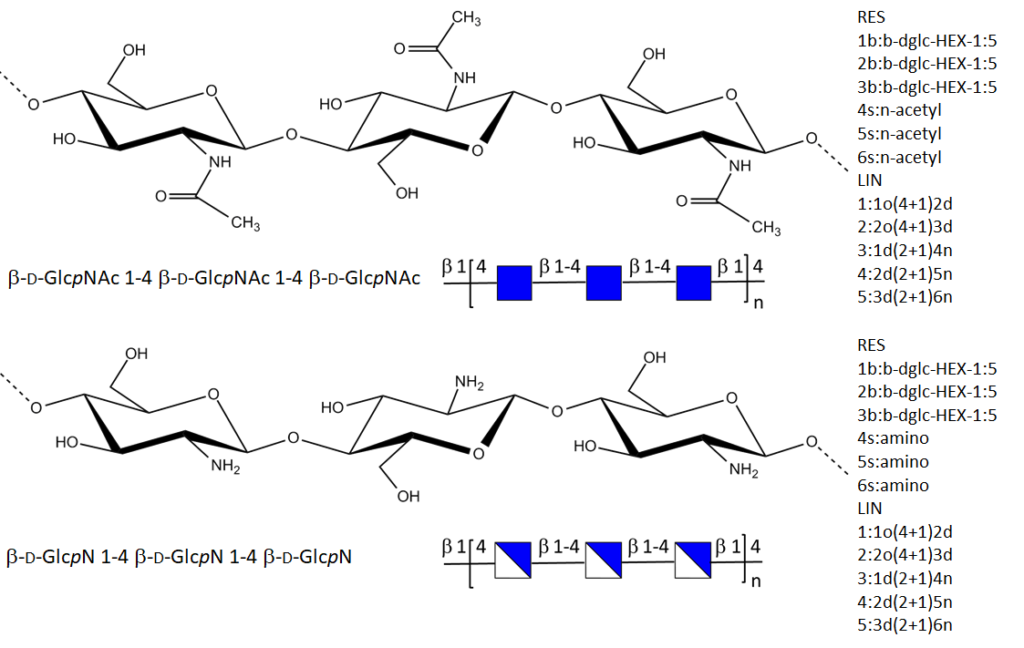
The descriptions and depictions of chitin and chitosan require several levels that are linked to the carbohydrate nature and the polymeric nature of these polysaccharides. As regards to their chemical compositions, it is a common practice to use IUPAC nomenclature, in the condensed form. (McNaught, 1977) A perspective drawing of the ring offers a simplified model […]
Chitins and Chitosan in the Solid State
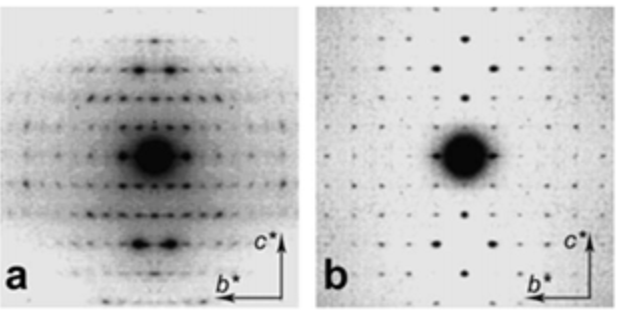
6.1. Crystallography of Chitins The availability of chitin in the form of a solid polycrystalline material made it a substrate of choice for X-ray diffraction studies, first on powders and later on oriented fibrillar arrangements. Clark and Smith were the first to make crystal studies of chitin and chitosan using X-ray diffraction.Clark & Smith, 1937 The powder […]
Fraction and Pattern of Acetylation
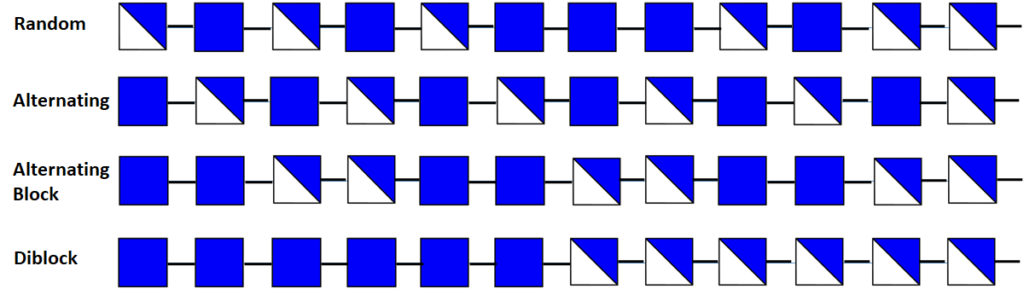
Chitosan is the collective name for a group of fully and partially deacetylated chitins. Their fraction of acetylation FA influences the properties of chitosan, (also described as Degree of Acetylation (DA) The pattern of acetylation (PA) and their degree of polymerization. These parameters influence the physicochemical properties strongly from which the solubility in acidic conditions […]
Chitin & Chitosan : Solubiity
8.1. Solubility of ChitinChitin occurs naturally partially deacetylated (with a low content of glucosamine units), depending on the sourceMathur & Narang, 1990nevertheless, both α and β forms are insoluble in all the usual solvents, despite natural variations in crystallinity. The insolubility is a major problem that confronts the development of processing and uses of chitin. […]
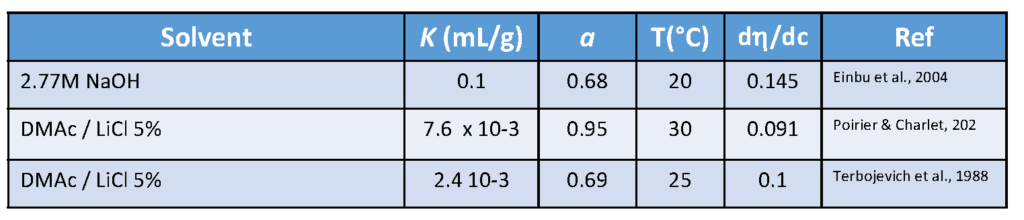
Chitin & Chitosan: Molecular Weight, Persistence Length
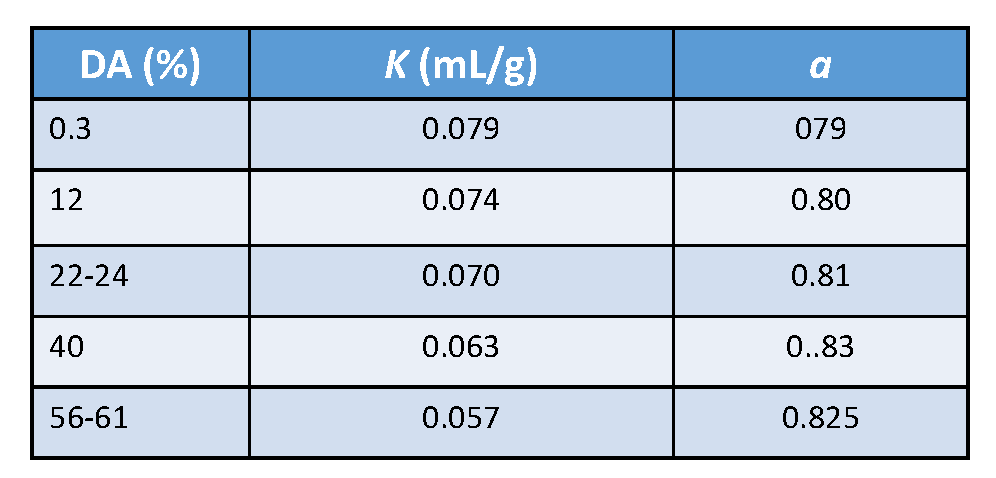
9.1. Molecular Weight. A complete analysis of the molecular weight distribution by SEC using triple detection Brugnerotto et al., 2001 (viscosity, concentration, molecular weight) was reported. The samples under investigation were heterogeneous chitosans : some were from commercial sources after solid-state treatment ; others were some homogeneous chitosans of different molecular weights obtained by re-acetylation of a highly deacetylated chitosan.Roberts […]
Chitosan : Complex Formation

10.1. Complex formation with metalsChitosan exhibits good complexing ability throughout the involvement of the –NH2 groups along the chain, in specific interactions with metals. Many articles have been devoted to the formation of complexes for the recovery of heavy metals from various wastewaters.Muzarelli, 1973 A mechanism for complex formation with copper at pH>5, was proposedDomard, 1987 (in agreement […]
Chitin and Chitosan Derivatives
11.1. Chitin Derivatives Chitin displays properties such as biodegradability, and non-toxicity, which form the basis of many applications in bio-medicine, pharmacy, biotechnology, food industry, nanotechnology.Arguelles-Monal et al., 2018. Due to the analogy between cellulose and chitin, reactions are often performed in similar experimental conditions. For highly acetylated chitin, reactions allowing to generate chitin derivatives involve the […]
Some Applications of Chitin and Chitosan
The high interest in medical applications of chitosan and some of its derivatives are readily recognized. The cationic character of chitosan is unique : it is the only pseudo-natural cationic polymer. Its film-forming properties and biological activity invite new applications. 12.1. Chitin-based MaterialsChitin has low toxicity and is inert in the gastrointestinal tract of mammals. It […]
References
A SERIES OF BOOKS ON CHITIN & CHITOSAN REFERENCES
Glyco Cyber Space
Glycoscience is a rapidly developing and emerging scientific discipline. Like many other scientific disciplines, glycoscience is adapting to the exciting rise of accessible scientific data, which now impacts research and modifies its practice. The accumulation of information along with the development of enabling technologies has laid the foundation of a rich computational toolbox tailored for […]
Introduction
Two recently published monographs, “A road-map for Glycoscience in Europe” and “Transforming Glycoscience : A Roadmap for the future” published respectively under the auspices of the European Science Foundation and the National Academies USA, identified a selected number of goals. One of particular importance was the need for the “establishment of long-term databases and bioinformatics and […]
Glycomics

Carbohydrates are a class of molecules utterly crucial for the assembly of complex multicellular organisms which requires interactions among cells. Beside the mediator role in cell-cell, cell-matrix, and cell-molecule interactions, glycans play an essential function in host-pathogen interaction. This is not surprising since all types of cell in nature are covered by a glycocalyx, a […]
Data Integration
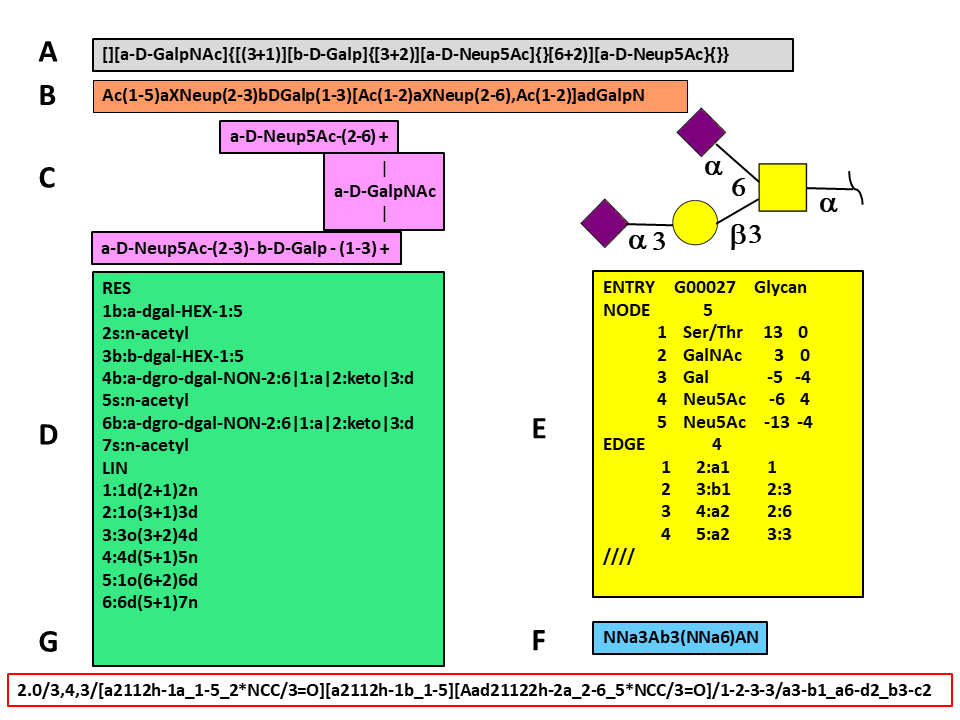
Data integration in glycomics This section presents the steps needed to integrate biological data within glycomics and with other “omics”. Glycan formats Glycans are inherently more complex than nucleic acids and proteins, so defining a format to store the molecular information correctly is not a trivial problem. The complexity of the glycans resides in their […]
Web Developments
Web developments The integration of multiple data source is critical to have a complete overview of a living system. Then, once data are correctly combined, it is essential to develop software applications which allow the exploration of the integrated data sources and the design new hypotheses. Thus, besides data integration, software development represents another critical […]
Overviews: Tools & Databases
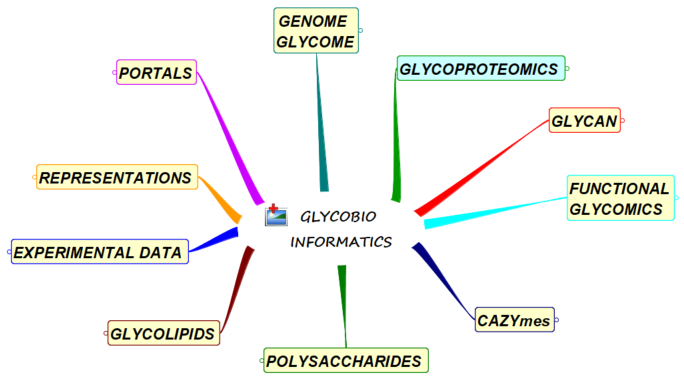
With the aim of facilitating glycoscience research, we have identified the tools and databases that are freely available on the internet and are regularly updated and improved. They have been clustered according to the major fields of applications: : Portals ; Genome and Glycome ; Representations ; Experimental Results ; Glycan ; Glycoproteomics ; Functional […]
Representations
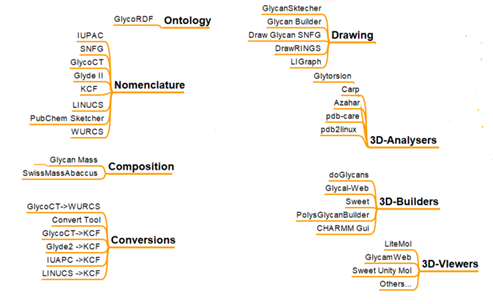
NOMENCLATURE CONVERSION : Utilities COMPOSITION DRAWING 3D-ANALYSERS 3D-BUILDERS 3D-VIEWERS
Experimental Results
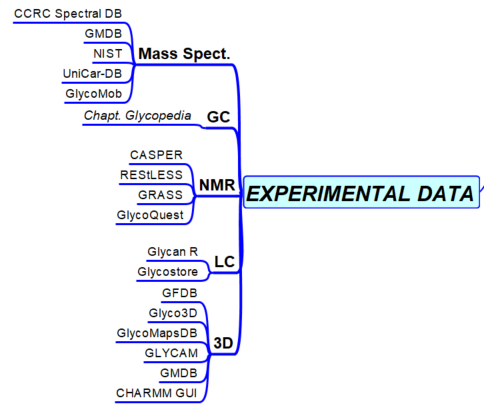
MASS SPECTROMETRY GAS CHROMATOGRAPHY LIQUID CHROMATOGRAPHY NMR 3D STRUCTURES
GlycoProteomics
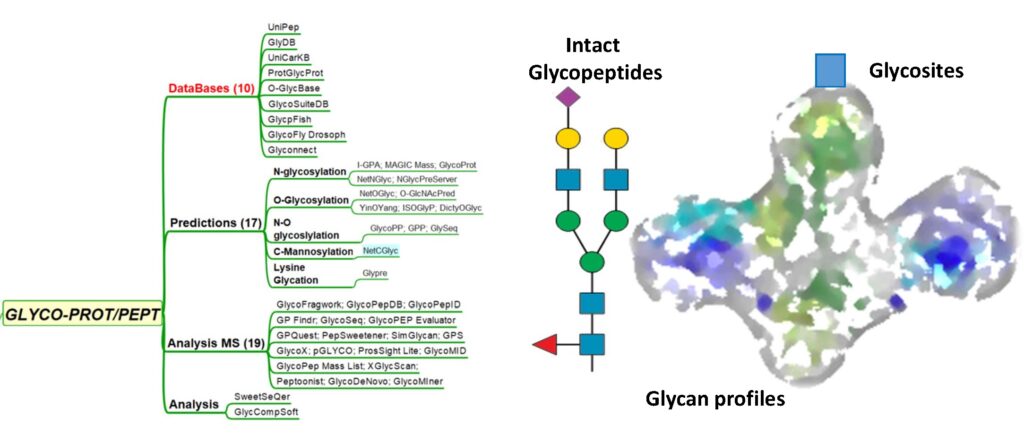
DATABASES PREDICTIONSN-Glycosylation O-Glycosylation N-O-Glycosylation C-Mannosylation Lysine Glycation ANALYSIS ANALYSIS MS
Glycans
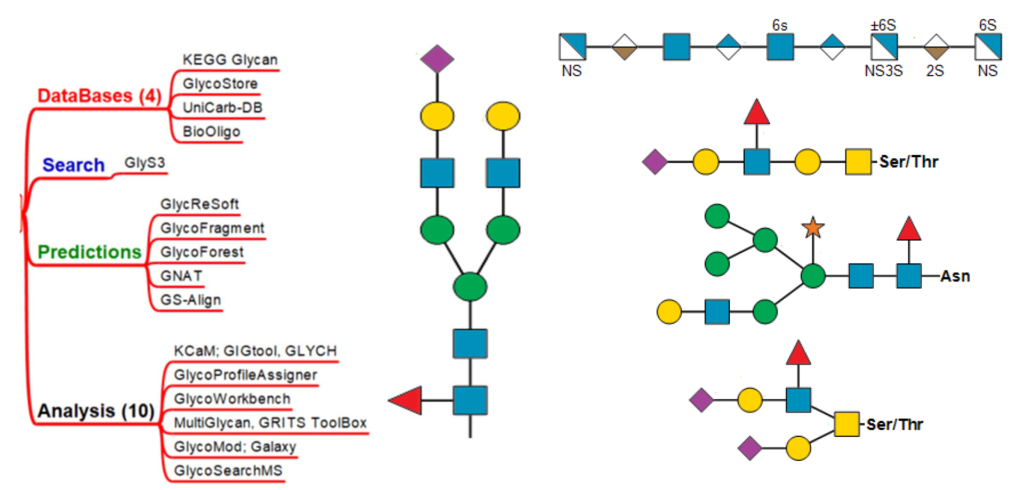
DATABASES SEARCH PREDICTIONS ANALYSIS
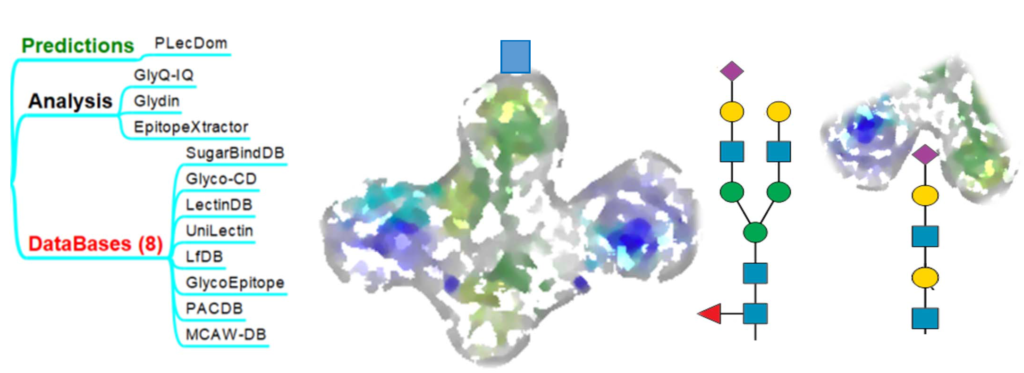
Functional Glycomics
DATABASES PREDICTIONS ANALYSIS
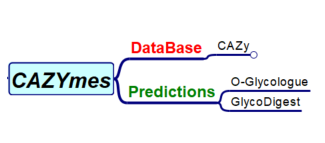
CAZYmes
DATABASES PREDICTIONS
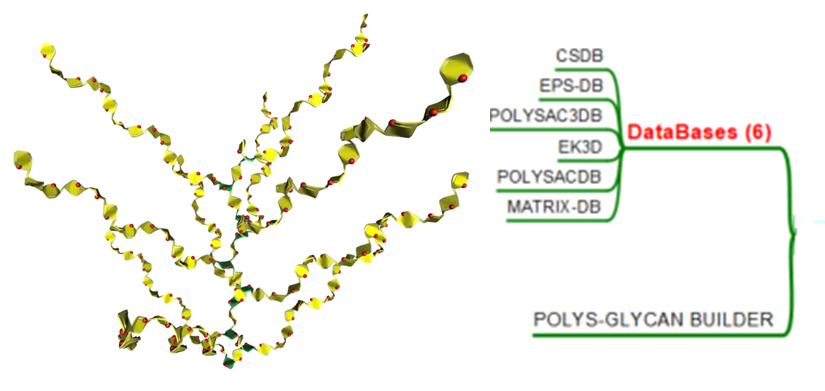
Polysaccharides
Glycolipids
Integrative Tools in Practice
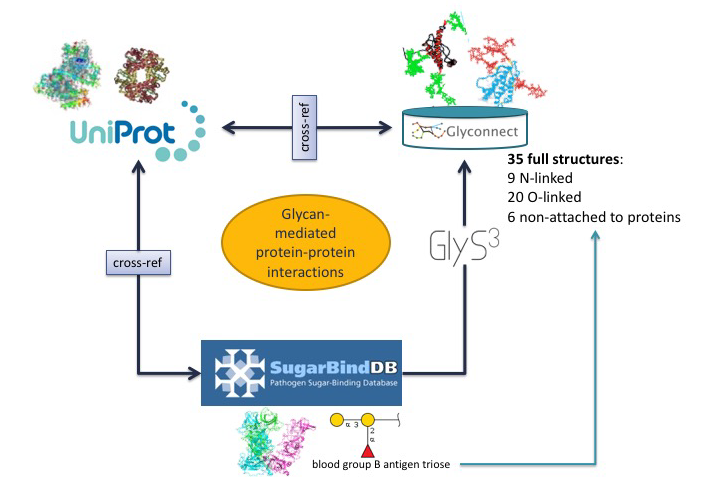
The collection of tools and databases described in detail in the previous sections can be grouped into two categories. They can either be dedicated to solving a specific question or be used in an integrative way in several applications. In the context of exploring and understanding the biological functions where glycans are involved, toolboxes are […]
Annex I: Data Integration
Data integration is one of the main challenges in bioinformatics. Although the term “data integration” often appears in research, especially in life science, a consolidated definition is still missing. We report here several definitions of “data integration” which will help in understanding the central concept and how it evolved. In the beginning, the “data integration” […]
Annex II: Data Integration Strategies
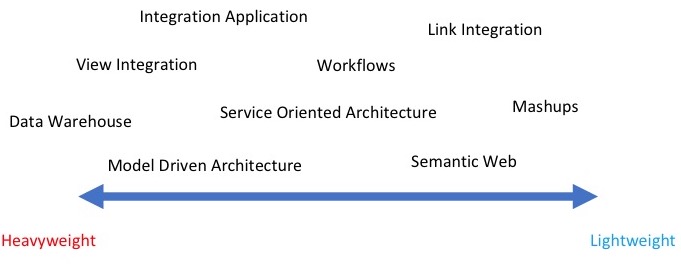
In the last 20 years, software developers have explored a wide variety of methodologies for data integration. Each of this methodology has a technology of reference and uses one or more touch-points. A touch-point is the logical connection between different data sources which make integration possible, for example, data values, names, ontology terms, keywords, etc. […]
Annex III: Data Integration in Bioinformatics
All data integration techniques presented in the previous paragraphs need touch points to be implemented. In bioinformatics, a diversity of efforts have been carried out to provide standards and, as a matter of fact, touch points across data sources. In this paragraph, we identify some areas of study which are crucial to enforce standardisation and […]
References
The Authors
Davide Alocci Trained as a Computer engineer at the University of Siena, I can define myself as a technology enthusiast. During an internship for the Master’s thesis, I discovered Bioinformatics, and it was love at first sight. After taking my Master in Computer Engineering, I decided to leave Italy, and I ended up in Geneva. […]
Introduction
The immune system is commonly conceived as the set of cells and molecules which participate in a concerted and tuned response to foreign substances, through a process called the immune response. These foreign substances encompass microbes as well as macromolecules (e.g., proteins, polysaccharides) and small chemicals, which are perceived through molecular recognition events as “non-self.” The […]
Innate, Adaptative, Passive-Active Immunities
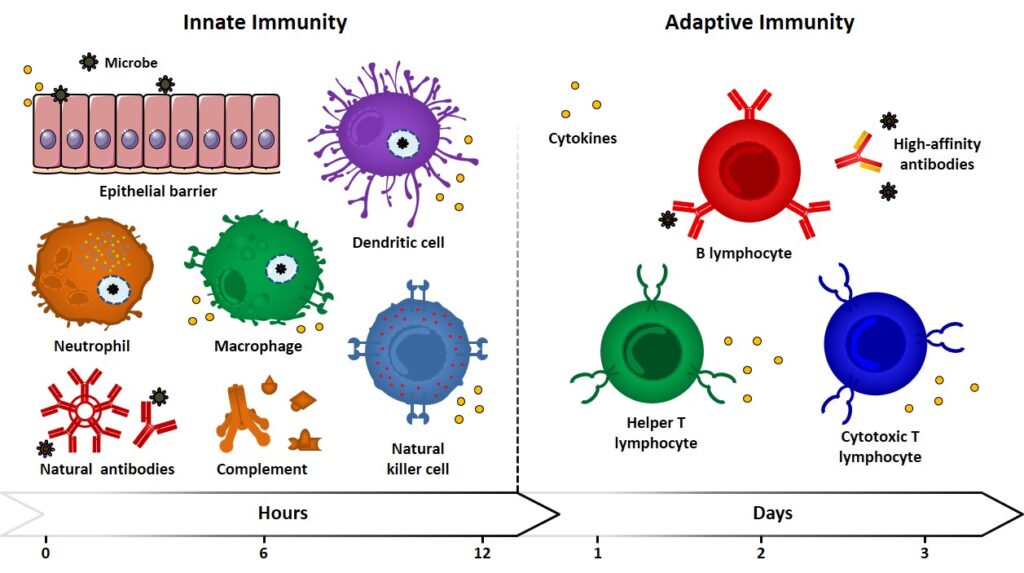
Innat immunity Innate (also referred to as natural or native) immunity operates in the early stage of the immune response by means of already existing mechanisms, which are reproduced every time the same kind of immunogen comes into play. Because of pathogen pressures during evolution, innate responses are directed against antigen motifs that are shared […]
Humoral adaptive immune responses
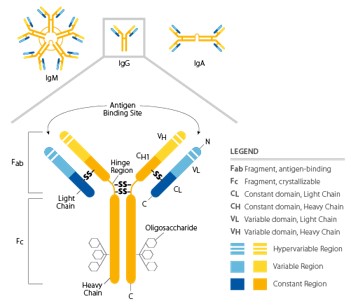
Humoral adaptive immune responses Humoral and cellular (or cell-mediated) adaptive responses are described by the different components involved in the immune-mediated clearance of pathogens such as microbes, toxins, infected cells or cancer cells. Humoral immunity exercises its functions by means of molecules found in the bloodstream and mucosal secretions, called antibodies (Abs) or immunoglobulins (Igs). […]
Cellular adaptive immune responses
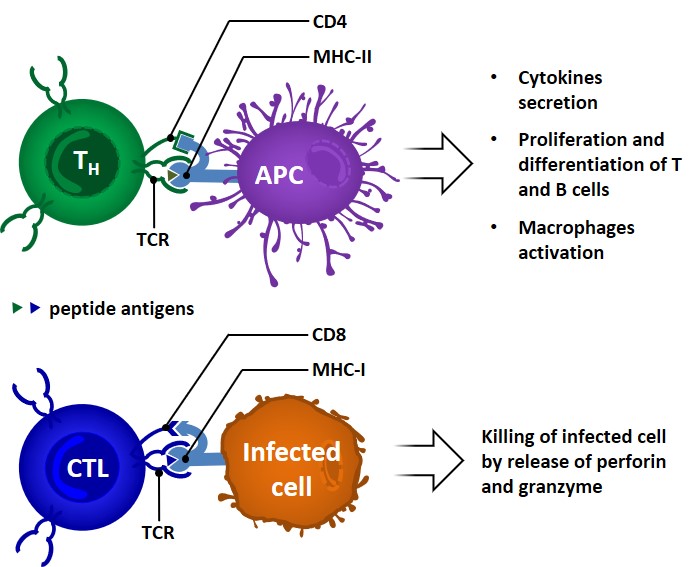
Cellular immunity is mediated by T lymphocytes (T cells). Intracellular microbes (eg. viruses) which are able to survive and proliferate inside phagocytes are not accessible to circulating antibodies; the evolutionary function of this type of adaptive response is meant to eradicate reservoirs of infection. Some T-cells also contribute to the clearance of extracellular microbes by […]
Antigen presentation
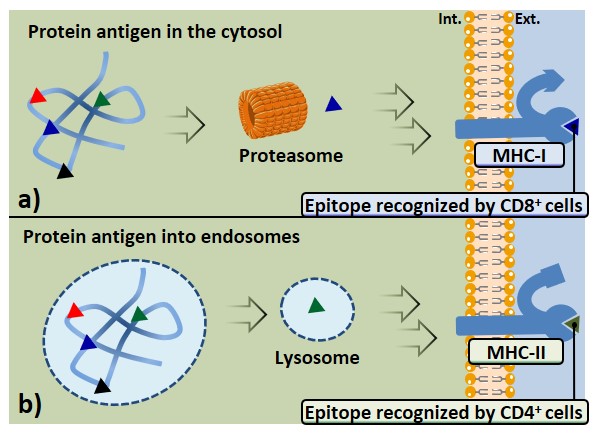
Although innate immunity reactions involve effective mechanisms able to control and even eradicate threats such as infections (not discussed in the present manuscript), many pathogens and transformed cells resist innate immunity. The attention is focused towards those factors which allow triggering more competent adaptive immune responses. As discussed earlier, the three main mechanisms of adaptive […]
Interplay between innate and adaptive systems
As mentioned above, T-cell’s functions are dependent on their physical interaction with other cells: they need to recognize antigens associated with MHC molecules on the cell surface. However, the modulation of the signal is dictated by secondary interactions involving molecules induced in APCs as the consequence of a previous antigen encounter (co-stimulators), and by the […]
Cell activation
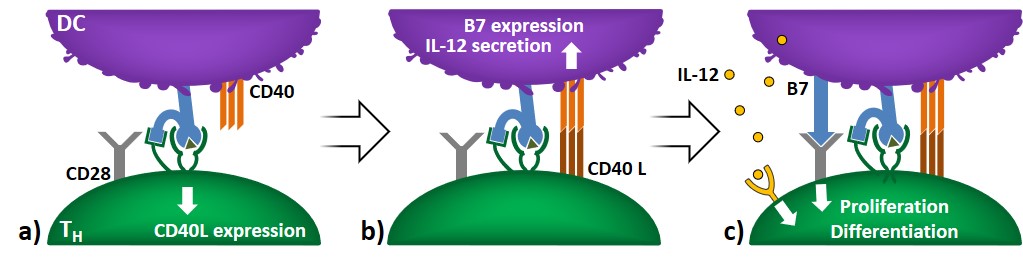
TH cell activation and licensing The first event in helper T-cell activation involves DC-mediated antigen recognition (with or without B7), which causes the expression of co-stimulatory CD40L on TH cells (Figure 7a). The binding of CD40L to CD40 increases the expression of B7 in DCs and stimulates cytokine secretion (e.g. IL-12). This causes the activation […]
Conclusions
The scope of this chapter was to present the main aspects of the immune response. They are summarized as follows : (i) Specificity ensures the immune response to be directed against the targeted microbe/antigen ; (ii) Diversity enables the spreading of the immune response towards a large variety of antigens ; (iii) Memory increase the ability to fight […]
References
The author
Carlo Pifferi Carlo Pifferi obtained his Master’s degree in Pharmaceutical Chemistry and Technology, under the supervision of Prof. C. Nativi, Dr. S. Roelens, and Dr. O Francesconi at the University of Florence (Italy), focusing on supramolecular chemistry and organic synthesis of synthetic receptors for the molecular recognition of carbohydrates. He then attended the University of […]
Introduction to Blood Group
The ABO(H) system although it is probably the oldest and most typed for human blood group system, yet from many perspectives, it is still poorly understood. The antigens of this system are sugar chains (glycans), which are present in red cell membranes, but also exist on most other non-erythroid cells, as well as in tissues/secretory […]
A simplistic overview
The presence of a blood group A and B antigens on the outside of a red cell starts with the presence of a blood group A or B gene and a variety of different glycolipid and glycoprotein blood group H antigen precursors. When the gene is transcribed it results in a glycosyltransferase, which in the […]
ABO genetics
The ABO gene was first cloned in 1990 and is located on the long arm of chromosome 9q34 Ferguson-Smith et al.,1976Yamamoto, et al., 1990and consists of seven exons and six introns, covering approximately 20 kilobase pairs from the initiation to the stop codon White, T., et al.,1995Bennett, E.P., et al.,1995 Yamamoto, 1995. The nucleotide sequence of the coding […]
Glycosyltransferase, a short brief
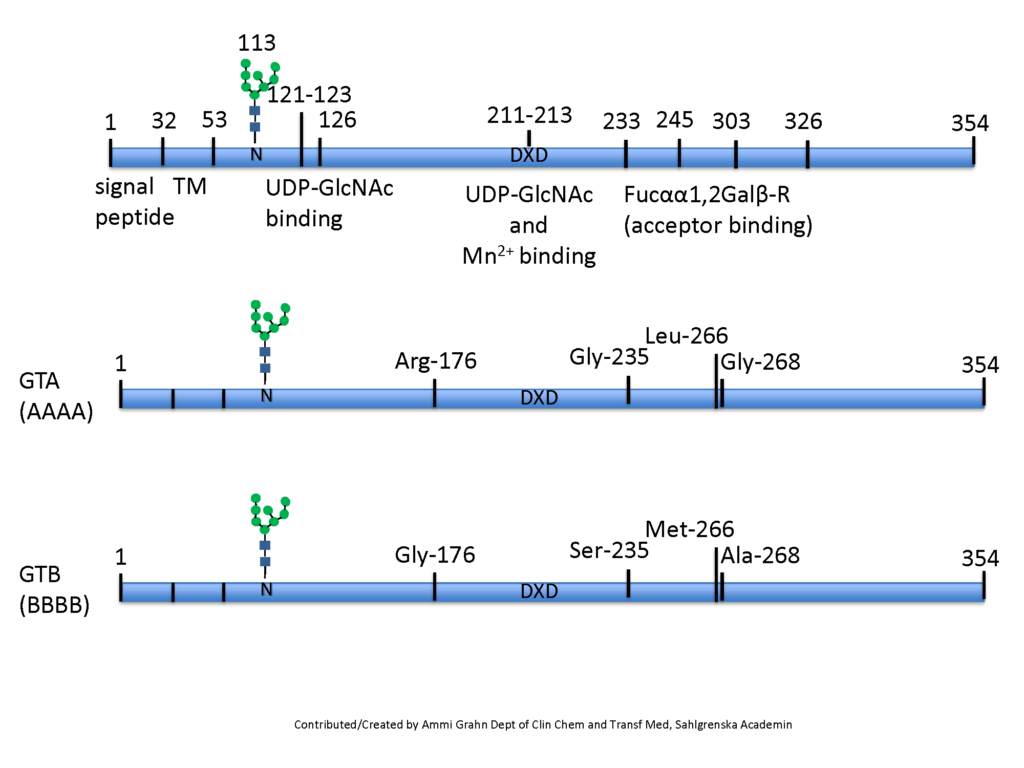
Glycosyltransferases are type II membrane proteins, with a short N-terminal cytoplasmic tail, a hydrophobic transmembrane region (TM) and the C-terminal catalytic domain in the lumen of the endoplasmic reticulum (ER) and Golgi. The cytoplasmic, transmembrane and stem regions of glycosyltransferases may specify their functional localization and stability within the Golgi Grabenhorst & Conradt, 1999. Reports have […]
Biosynthesis of ABO and related antigens
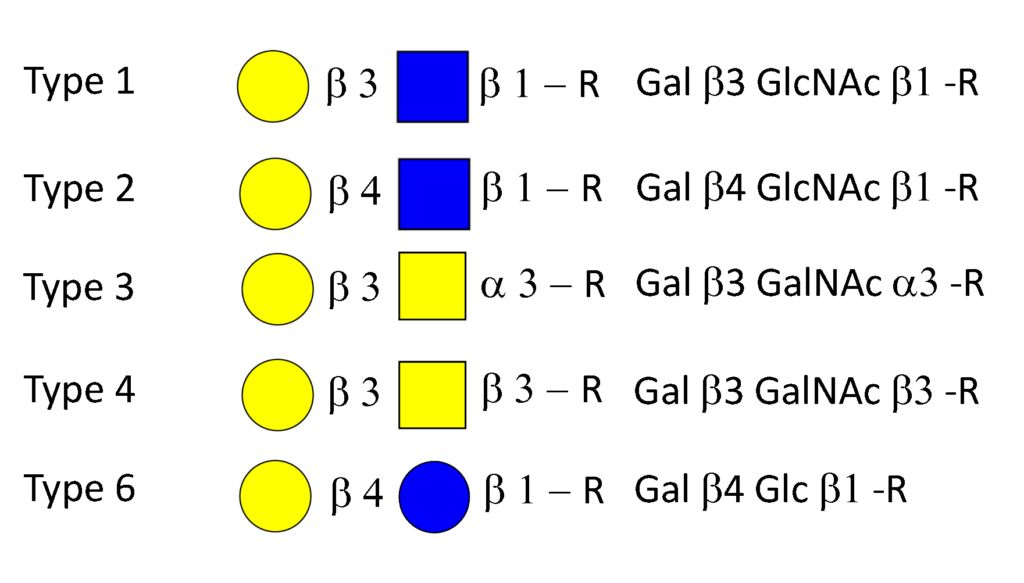
Synthesis of ABO antigens, like all carbohydrate antigens, is the result of a complex series of interactions between a range of different glycosyltransferases residing in the endoplasmic reticulum and Golgi apparatusColley, 1997Maccioni et al.,2011aMaccioni et al., 2011b. To form an antigen these glycosyltransferases must catalyse reactions in an orderly manner between a variety precursors and […]
Biosynthesis of ABO glycolipids
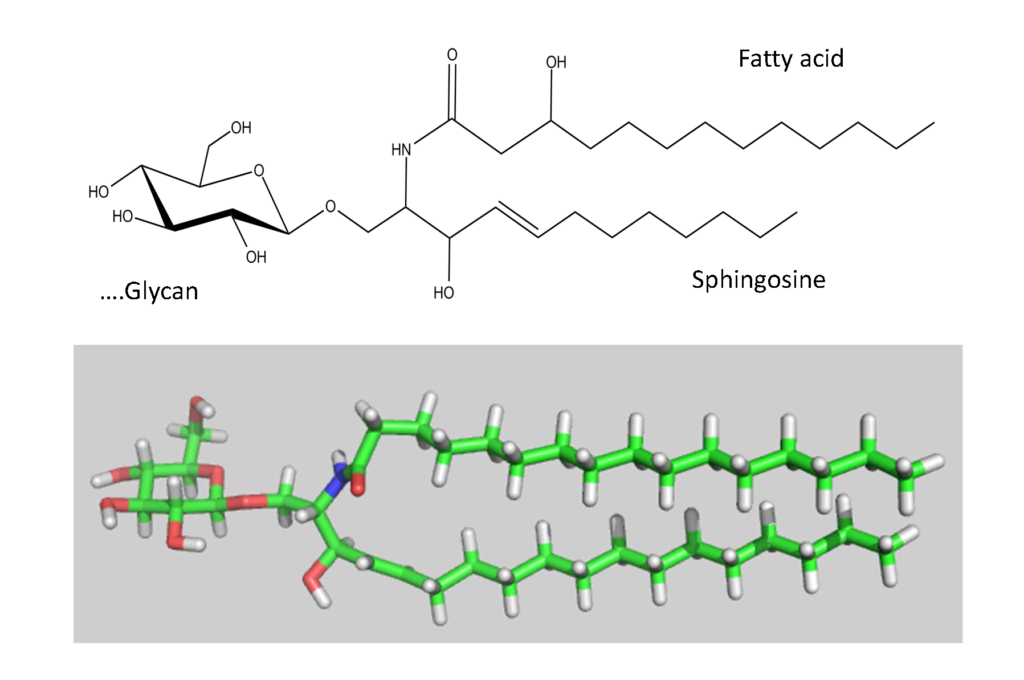
The biosynthesis of the ABO and all its related precursors such as H, I and P blood group system glycolipids (glycosphingolipids) all originate with glucose (Glcβ1) linked to a ceramide (Figure 5). A galactose residue in β4 linkage (Galβ4) forms lactosylceramide (Galβ4Glcβ1-Cer) (Figure 6). From this precursor two alternative pathways exist, one via lactotriaosylceramide towards […]
Biosynthesis of glycoproteins
The biosynthesis of glycoprotein based blood group A, B and H antigens relating to terminal glycosylation is similar to glycolipids in most aspects. However, biosynthetically blood group protein glycosylation starts very differently from glycolipids and usually involves two different pathways. The first starts with the linkage of a GalNAc residue to oxygen of a serine […]
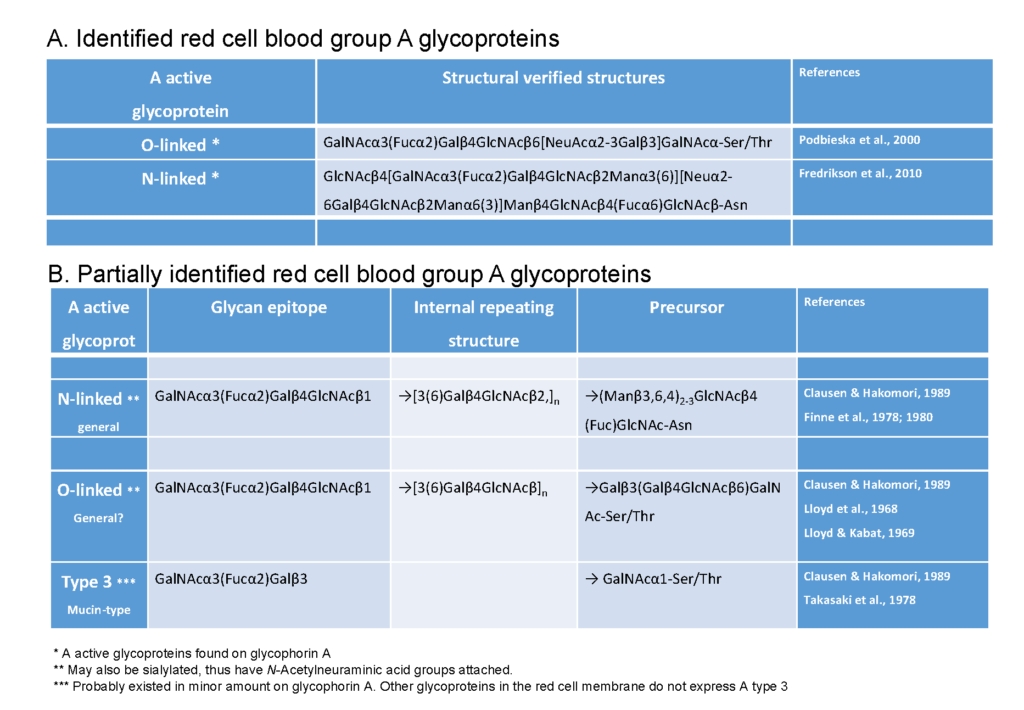
The antigen on the erythrocytes
The ABH antigens of red cells are carried by both glycoproteins and glycolipids on a variety of “peripheral cores” that influence their antigenicity Clausen & Hakomori, 1989Watkins et al., 1988. It should be noted that there is also a range of “internal core” structures on O-linked proteins using different terminology ; but these do not directly influence […]
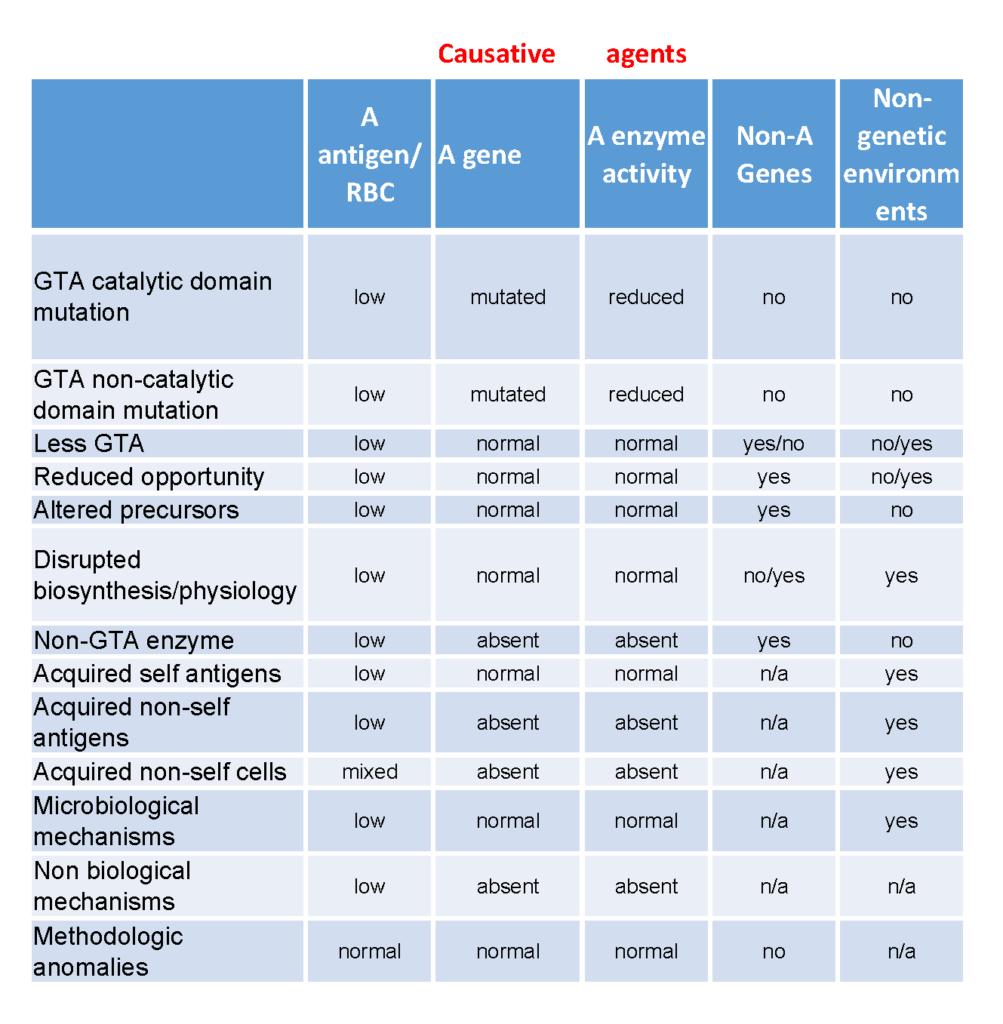
Influence of other systems on ABO antigens
In addition to those interactions with glycosyltransferases involved in biosynthesis described in previous chapters above (Figure 4), the effects of other blood group systems on ABO antigens deserve further mentioning. These ABO unrelated blood group systems can independently and cumulatively have consequences on ABO antigen expression either by competing for precursors and/or structurally altering the […]
ABO and phenotypes
Anti-ABO antibodiesA and -B antibodies are present in individuals lacking the specific ABO antigen.The principles described in this section of antibodies against blood group A antigen are in most cases also applicable to blood group B antigen. Polyclonal antibodies (PAb) associated with blood group AAnti-B appears always to be associated with all healthy individuals with […]
References
Ando, S., et al., A novel pentaglycosyl ceramide containing di-beta-N-acetylgalactos-aminyl residue (Para-Forssman glycolipid) isolated from human erythrocyte membrane. Adv Exp Med Biol, 1982. 152 : p. 71-81. Anstee, D.J., Blood group-active surface molecules of the human red blood cell. Vox Sang, 1990. 58(1) : p. 1-20. Bennett, E.P., et al., Genomic cloning of the human histo-blood group ABO locus. Biochem Biophys Res Commun, 1995. 211(1) : p. […]
The authors
Lola Svensson Lola Svensson received her training as a biomedical scientist in Clinical Chemistry and Transfusion Medicine from the Univerisity of Gothenburg, Sweden. She defended her PhD in Clinical Chemistry on the ”Chemical basis of ABO subgroups. Insights into blood group A subtypes revealed by glycolipid analysis”. The PhD studies were focused on the chemical […]
Introduction
(A hard-copy is available at :http://www.glycopedia.eu/IMG/pdf/the_plant_cell_walls.pdf) Observing a section of cork with a microscope that he had himself devised, Robert Hooke, discovered that the tissue had a honeycomb-like structure made of small spaces contained by walls, that he coined cells (from the latin word cellula, small compartment). He recorded his observations in his famous book Micrographia […]
Plurality of Cell Wall Types
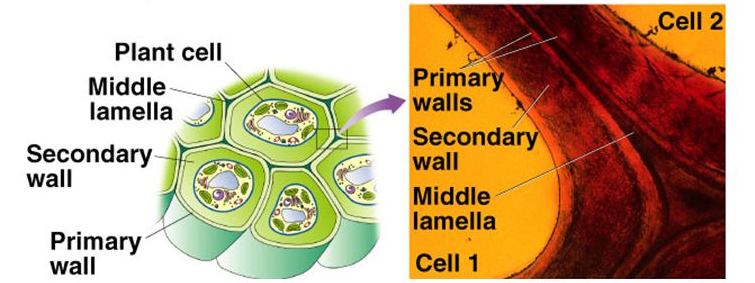
The constituents of the plant cell walls vary depending on the stage of development and the type of tissue. In fact, when analyzing the structure-function relationship, one should always consider the plurality of plant cell types (ca 35 different types in plants, Cosgrove, 2005) with characteristic walls since their composition and macromolecular organization are adapted to a […]
Polysaccharide Diversity
During photosynthesis plants fix the carbon from carbon dioxide into sugars which are incorporated into complex polysaccharides. This process makes plant cell walls a terminal carbon sink and constitutes the dominant carbon sequestration system on Earth. All plant cell walls share in common the characteristic of having a complex composite structure organized on a cellulose […]
Cellulose, Pectins,…..
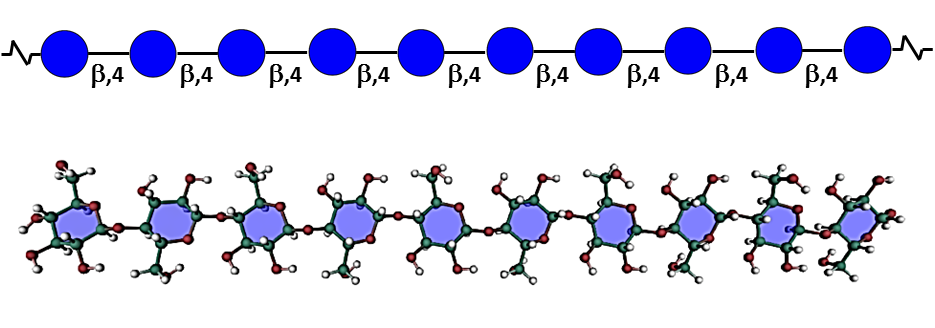
Cellulose This is the most abundant natural polymer on earth. Cellulose accounts for 15–30 % dry weight of the primary cell wall and 20-35 of the secondary walls. Its apparently simple primary structure is that of a homopolymer consisting of long linear chains of β-(1-4)-linked D-glucopyranose residues associated into the repeating disaccharide unit of cellobiose. The […]
Primary Wall Hemicelluloses
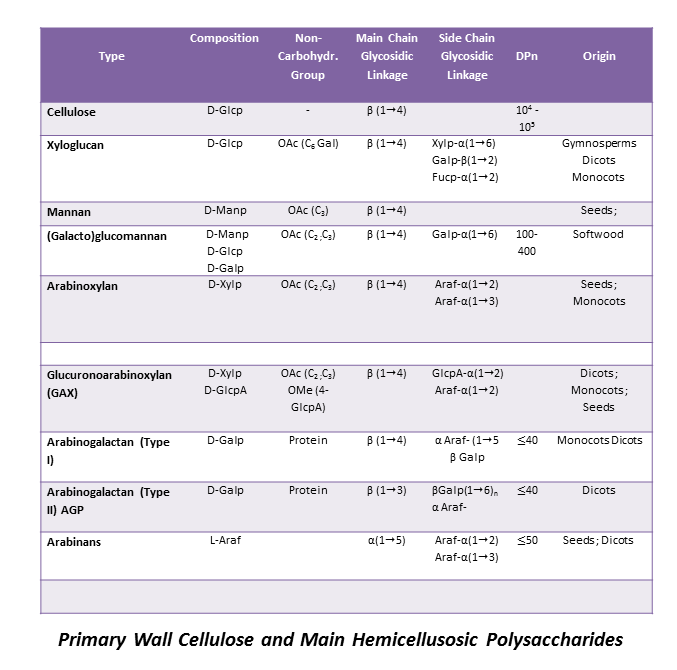
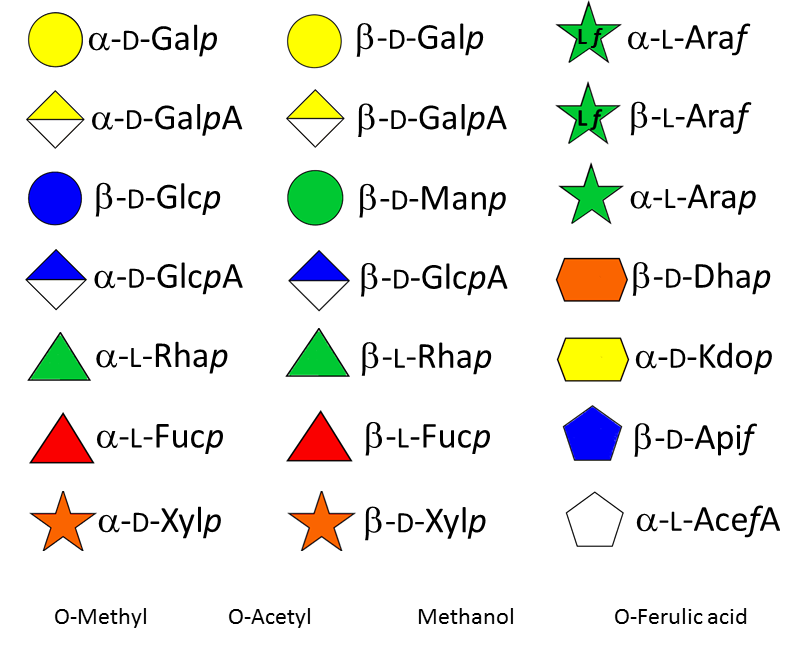
Xyloglucan (XG) They consist of a β-(1-4)-linked D-glucopyranosyl backbone carrying various side chains of D-xylopyranosyl-α-1-, D-galactopyranosyl-β-(1-2)-D-xylopyranosyl-α-1- , L-arabinofuranosyl-(1-2)-D-xylopyranosyl-α-1-, and (except in grasses) L-fucopyranosyl-α-(1-2)-D-galactopyranosyl-β-(1-2)-D-xylopyranosyl-α-1-.Fry 1989 The cellulose-like backbone is responsible for the strong and particular interaction between XGs and cellulose microfibrils. An interesting feature of this typical primary wall hemicellulose is that a certain regularity of distribution […]
Secondary Wall Hemicelluloses
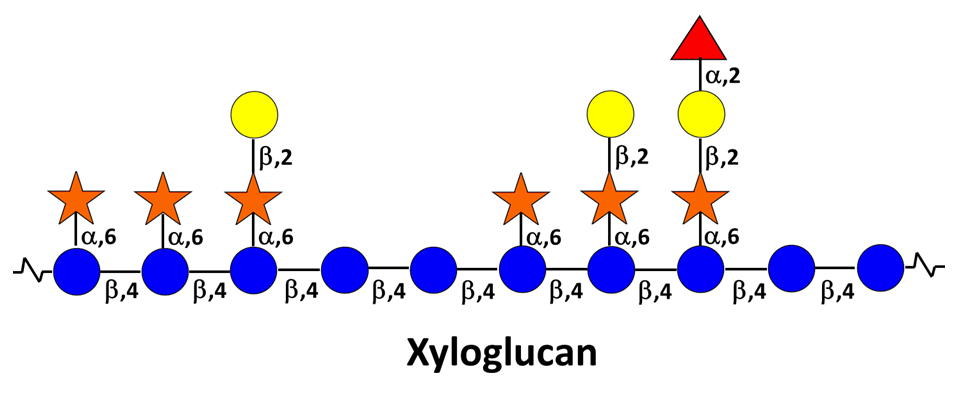
The thick secondary walls are laid down only after the cambial cells have stopped enlarging. They have a layered structure in which the layers are conventionally denoted S1, S2, etc., in the order in which they are laid down. harris 2006 Some cell types, such as parenchyma cells of the cotyledons and the endosperms of […]
The Plant Cell Walls are Complex Nanocomposites of Polysaccharides

Whether primary or secondary, the plant cell walls are built on the cellulose network organized around the cellulose microfibril unit (MF). Depending on the developmental stage, the tissue type, and the cell wall layer, cellulose microfibrils (MFs) are differentially embedded in pectic polysaccharides, hemicelluloses and lignin. Many fundamental biological, physical, mechanical and chemical properties of […]
Models of Plant Cell Wall Architecture
The plant cell walls can be viewed as a supramolecular complex of polymers intertwined in an organized manner. How the various and complex polysaccharides, proteins, phenolic compounds and inorganic ions are organized and interact to form the structural entity of the plant cell wall remains in great part unknown. Several models have proposed tentative structures […]
References
The Authors
Jean-Paul Joseleau Jean-Paul Joseleau received his education in chemistry at the University of Grenoble (1959-1962), and obtained a Ph.D. degree in organic Chemistry in 1965 at the University of Grenoble on lignin biosynthesis under the supervision of Professor F. Barnoud and Professor T. Higuchi (Japan). He was introduced to glycoscience research by Professor G.G.S. Dutton […]
Introduction
Bacteria convey unique biological structures that partly relate to a well-developped cell-wall structure structure. Thanks to the relative simplicty of bacteria (as opposed to larger organisms) many investigations of the cell-wall structure have been performed and shown to be the source of the understanding of many biochemical processes that have been subsequently shown to be […]
Main Features of the Bacterial Cell Wall
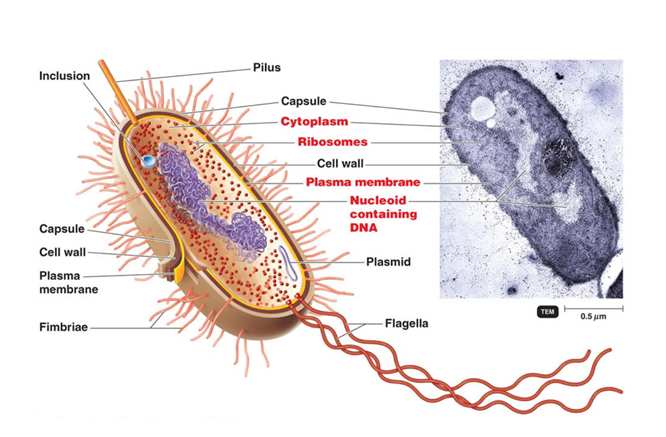
Bacteria are unicellular organisms, which are spread in a wide variety of environments, from our gut Modi et al., 2014 to the deepest layers of the oceanic crust Mason et al., 2010. These organisms can adopt a wide variety of shapes, from cocci (the spherical Staphylococcus aureus), to bacilli (the rod-like Escherichia coli) or even spirilla (the helically coiled Helicobacter pylori). […]
The Bacterial Cell Wall
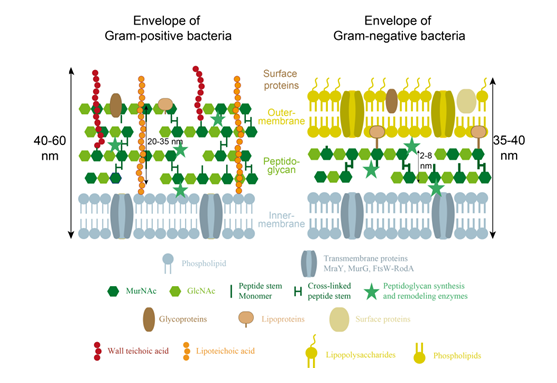
In 1884, the Danish bacteriologist, Hans Christian Gram, while trying to set up a protocol to stain bacteria for observation under the microscope, developed a technique, which became fundamental to discriminate bacteria according to the composition of their cell wall. Heat-fixed bacterial cells are first treated with a purple dye, gentian violet, which penetrates through […]
Macroscopic Structure of the Peptidoglycan Layer
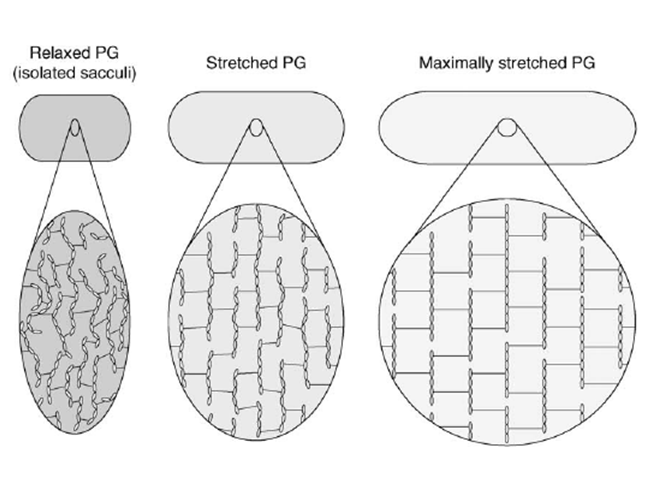
Although the presence of a peptidoglycan network in bacterial cell walls is well documented, the mechanical properties and architecture of the massive murein sacculus ( 3 x 109 Da, a repeat of approximately 3.5 x 106 moles of monomeric motifs in the Gram-negative E. coli as measured by Wientjes et al., 1991 are still poorly understood. […]
Peptidoglycan Molecular Structure
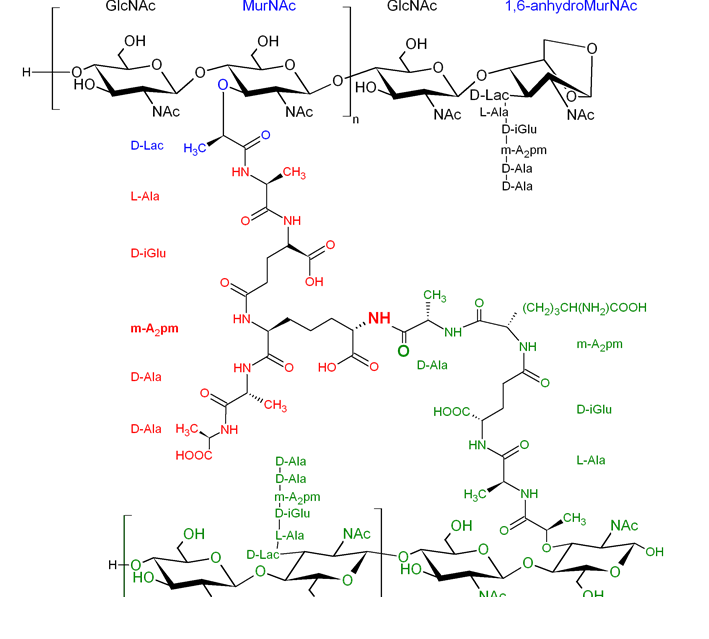
Chemical composition In all bacteria, the glycan strands of peptidoglycan are composed of alternating β-1,4-linked N-acetylglucosamine (GlcNAc) and N-acetylmuramic acid (MurNAc, a variant of GlcNAc with a D-lactate attached to the C-3 by an ether bond) as shown in Figure 8. Generally, the obtained strands are terminated at the reducing end by a 1,6-anhydroMurNAc residue, […]
References
The Authors
Nicolas Jean Nicolas Jean obtained a Bachelor Degree in Biology and a Master Degree in Structural Biology and Biochemistry from Université Joseph Fourier, Grenoble, in June 2010 and 2012, respectively. He joined the team of Jean-Pierre Simorre at IBS in January 2012 for his master thesis, where he started to work on the structural characterization […]
Presentation

Under the name “Sweet Existence” Karen Barber presented a vibrant, colourful collection of abstract paintings on the occasion of a scientific event organized by IBCarb in July 2015, in Manchester. IBCarb – Glycoscience Tools for Biotechnology & Bioenergy, is a BBSRC funded network in Biotechnology and Bioenergy. As a growing network of academics and industrialists […]
The Gallery

At the beginning Pulse I Pulse II Pulse III, IV,V,VI The Extra Cellular Matrix Monosaccharides & Polysaccharides The Endoplasmic Reticulum The Golgi Apparatus Gold Plate I Gold Plate II Gold Plate III Miracle of Life I Miracle of Life II Power of Light Vascular Bundle Starch Brillance Blue Green Algae I Blue Green Algae II
The Author
Karen Barber is primarily interested in celebration. Bold, provocative and daring, Karen’s vibrant work warms the senses to celebrate the joy of life and the beauty of colour and luxurious materials. Karen specialises in producing cell and science inspired work. Karen aims to convey joy, hope and energy through her range of paintings, screen-prints and […]
Introduction
Carbohydrates, in the form of mono-, oligo-, poly-saccharides, glycopeptides, glycolipids, glycosaminoglycans, proteoglycans, or other glycoconjugates, constitute the most abundant and complex family of bio(macro)molecules. They have long been known to participate in many biological processes. From their simplest to their most complex forms, they are divided into several categories, covered by the generic term of glycans. […]
General Concepts Related to Monosaccharide Analysis.
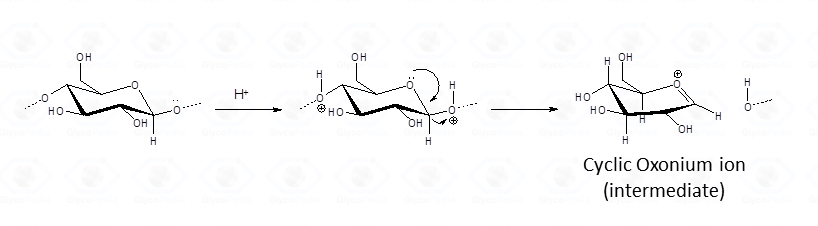
The coupling between Gas Chromatography (GC) and Mass Spectrometry is a powerful analytical method, having a very good detection limit, and prone to automation. Such an analysis requires the compounds to be volatile, a condition which is fulfilled neither by glycan nor even by single monosaccharides. As a consequence, chemical manipulations of the samples are […]
GC-MS Determination of Sugar Composition
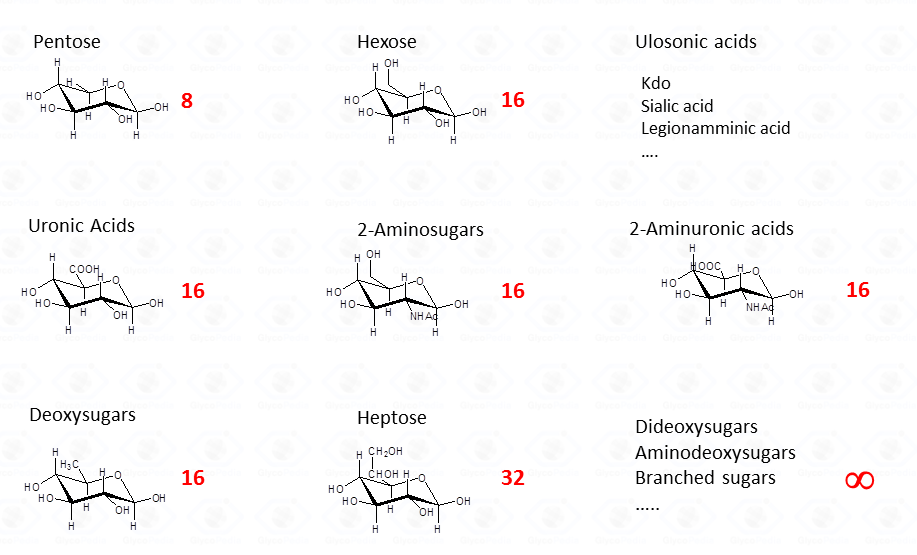
Many types of monosaccharides exist in nature, and they can be divided into different categories according to: (i) their nature (neutral, acidic and basic), (ii) the number of constituting carbon atoms (pentose, hexose, heptose, etc), (iii) the number of hydroxyl groups replaced with an hydrogen atom (deoxy, dideoxy, etc.), (iv) the aldose or ketose nature. […]
Acetylated Alditols: Advantages & Limitations

The chemical approach to produce these derivatives follows the scheme described below and experimental details are availabe elsewhere [link to “Further Readings”]. This procedure is suitable for the detection of aldose and ketose monosaccharides, but it is limited to neutral or basic species, so that acidic monosaccharides cannot be detected, unless the reduction of the […]
Acetylated Methyl Glycosides: Advantages & Limitations
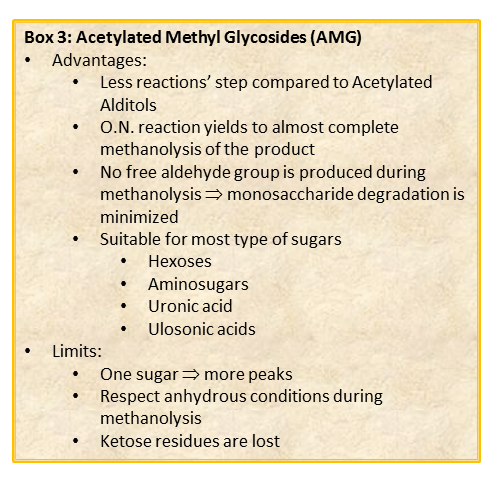
The chemical approach to produce these derivatives follows the scheme shown below shown, and experimental details can be found elsewhere [link to “Further Readings”]. Whereas this procedure is suitable for the detection of almost all type of monosaccharides (neutral or basic aldoses, uronic and ulosonic acids), it fails in the detection of ketoses, as fructose. […]
Partially Methylated Acetylated Alditols: Advantages & Limitations.

The chemical approach to produce these derivatives follows the scheme shown below. Experimental details can be found elsewhere [link to “Further Readings”]. In analogy with the approach discussed for the acetylated alditols derivatives, this procedure is well suited to aldose and ketose monosaccharides. However, it is limited to neutral or basic species. In the in […]
Acetylated Octyl Glycosides
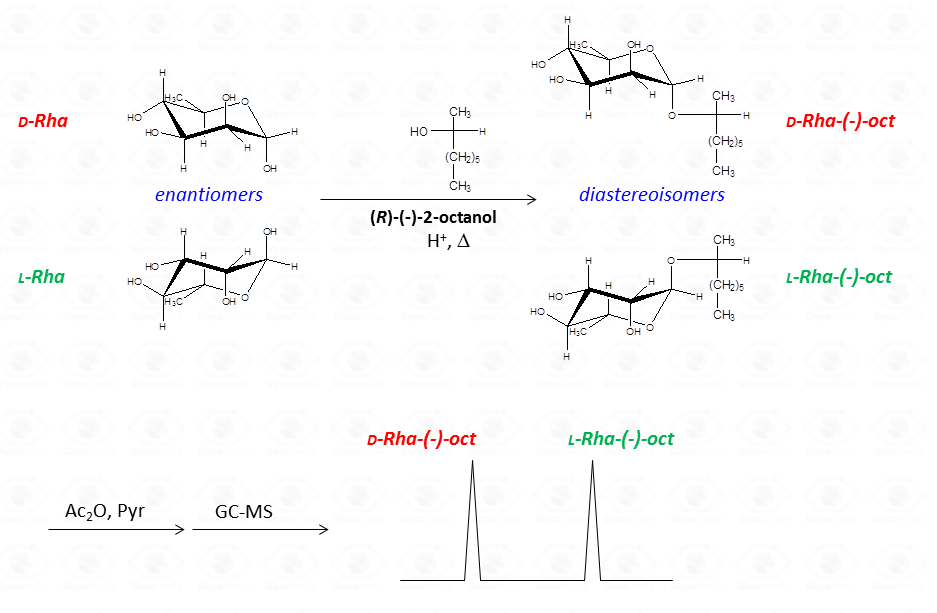
Analysis of these derivatives is a way to assess the absolute configuration of a monosaccharide unit. In nature, many monosaccharides such as rhamnose, fucose, or arabinose, occur with both D and L configurations. None of the three methods discussed so far, is capable of assessing the absolute configuration. For example in the case of acetylated […]
Further Reading
Experimental detail regarding the preparation of all the derivatives discussed in this review can be found in: De Castro C, Parrilli M, Holst O, Molinaro A. Microbe-Associated Molecular Patterns in Innate Immunity: Extraction and Chemical Analysis of Gram-Negative Bacterial Lipopolysaccharides. Methods Enzymol. 2010, 480, 89-115. Extended discussion of EI-MS spectra of all the derivatives discussed […]
The Author
Cristina de Castro Dr. Cristina de Castro graduated in 1992 with honors and November 1993, she started her Ph.D. in Chemistry, in the University of Napoli Federico II, focusing her attention on the structural determination of polysaccharides from phytopathogenic bacteria. After completing her PhD, she worked for 16 months in the Chemistry Laboratories of the […]
Annex
Introduction
Selectin Ligands and Related Structures on the Surface of the Infected Cell. Human herpesviruses are usually acquired early in life and are widely distributed in the population. A common feature of all human herpesviruses is that they persist in the host after the primary infection. Thus, the host immune system resolves the acute stage of […]
Herpesviruses
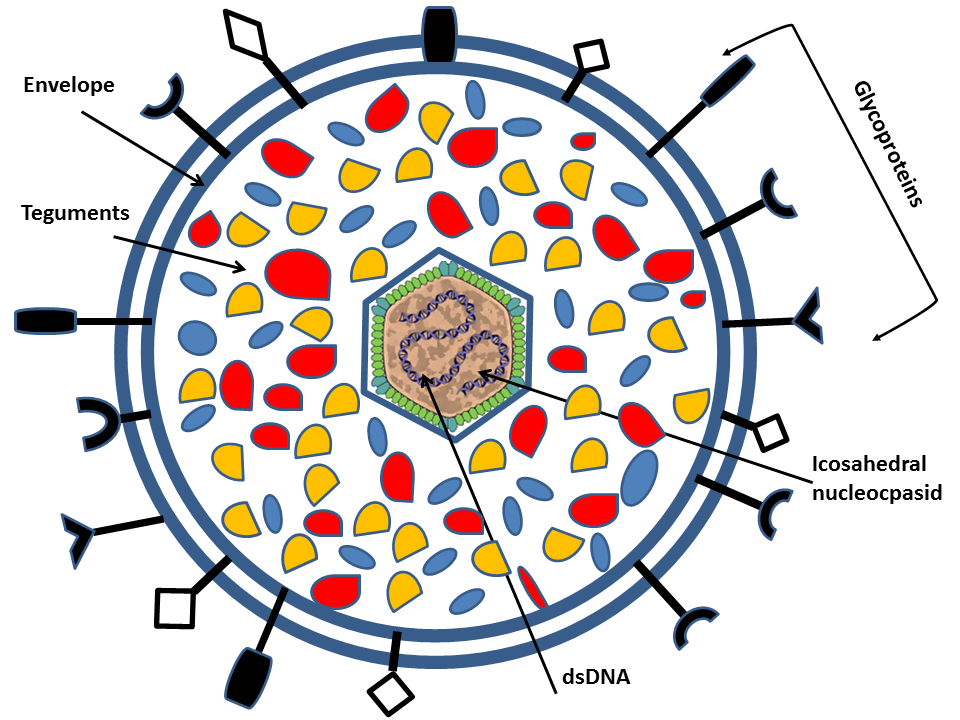
Human herpesviruses : Structure and basic properties Human herpesvirus is a family (Herpesviridae) of large enveloped double stranded DNA (dsDNA) viruses of which eight members have humans as the natural host. The herpesvirus particle has a diameter of about 200 nm and consists of an icosahedral nucleocapsid of 162 capsomers that is surrounded by a double […]
Normal and Pathological Colonization of Organs by Circulating Leukocytes
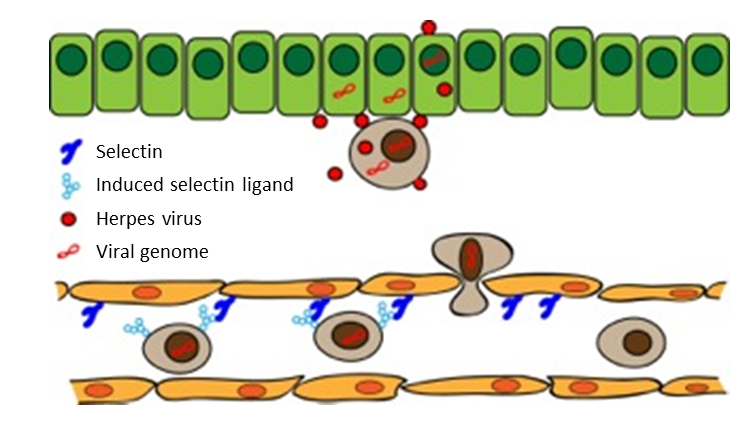
Licencing of leukocytes to pass the endothelial wall Occasionally, certain leukocytes of the blood stream have to cross the endothelial wall to perform tasks in adjacent tissue, i.e. to combat invading bacteria or viruses. However, the process by which circulating leukocytes may cross the endothelial wall to access adjacent tissue is strictly regulated, not least […]
Leukocyte Migration over the Endothelial Wall
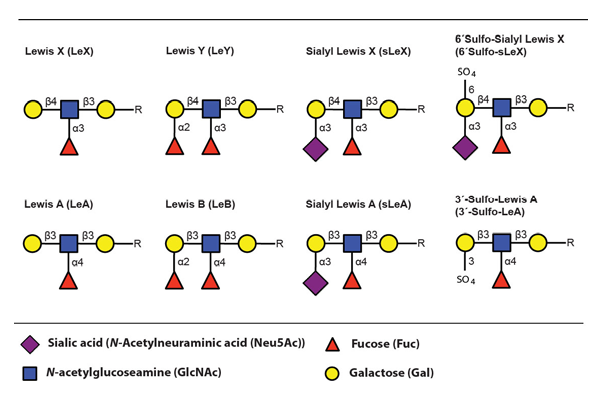
Selectin Ligands – Versatile Carbohydrate Epitopes. Selectins Selectins are carbohydrate-binding proteins that mediate reversible interactions with glycoconjugates enabling tethering and rolling of the leukocytes along the endothelium, a prerequisite for subsequent passage across the endothelial wall. (Zarbock et al., 2011 – Sperandio et al., 2009). E-selectin is expressed mainly on endothelial cells and P-selectin that can be […]
References
References Afshar-Kharghan, V., et al., Human polymorphism of P-selectin glycoprotein ligand 1 attributable to variable numbers of tandem decameric repeats in the mucinlike region. Blood, 2001. 97(10) : p. 3306-7. Allen, U. and J. Preiksaitis, Epstein-barr virus and posttransplant lymphoproliferative disorder in solid organ transplant recipients. Am J Transplant, 2009. 9 Suppl 4 : p. S87-96. Andersson-Ellstrom, A., B. Svennerholm, and L. Forssman, Prevalence […]
The Author
Rickard Nordén Graduated as a Master of Science (Molecular Biology) from University of Gothenburg in 2007. In 2008 he joined the Glycovirology group, headed by Professor Sigvard Olofsson, at the University of Gothenburg. The PhD project aimed at elucidating the mechanisms behind Herpesvirus manipulation of the cellular machinery for protein glycosylation. The work focused to […]
Editorial Introduction

Entitled “45 Years at Work with Glyco”, Prof. Hans Kamerling gave the opening lecture at the 13th European Training Course on Carbohydrates (Summer School Glycoscience) this April (2014) in Wageningen, The Netherlands. His lecture is a testimony to the evolution of glycoscience over 45 years of scientific endeavor, methodological and technological developments. The presentation includes […]
The Early Days.

From PhD to Post-Doctoral Stays
Glycosidic Linkage Analysis
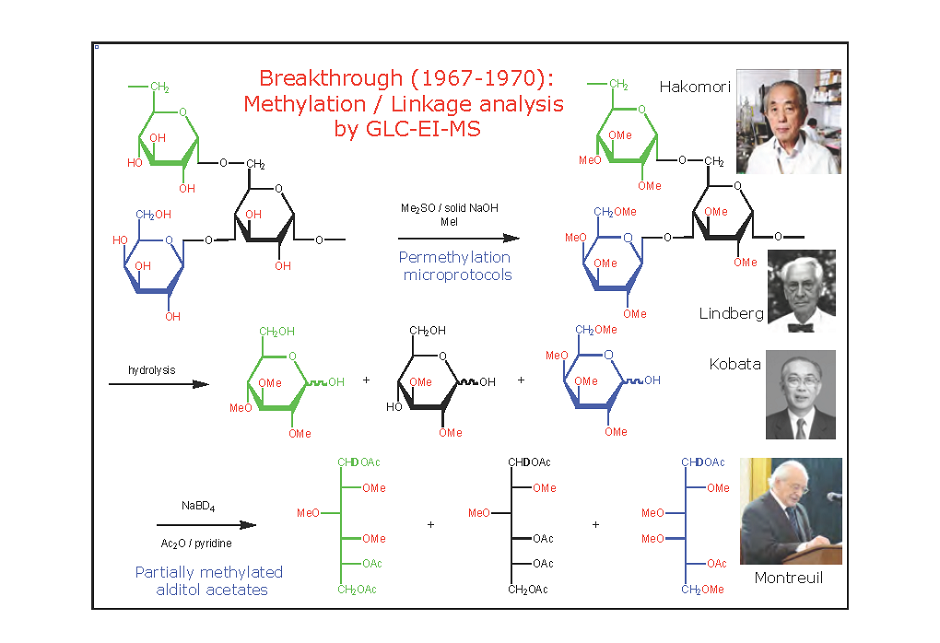
Methylation – Glycan -Polysaccharides
Absolute Configuration

From Fischer, Bijvoet ….
The Family of Sialic Acids
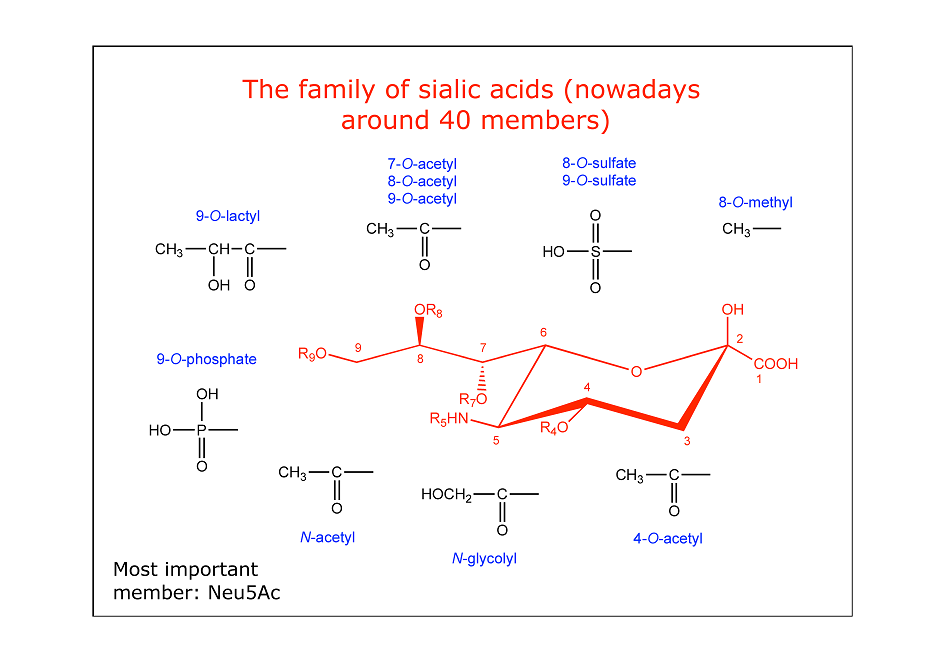
Oligosaccharide-based Vaccines
Carbohydrate-Carbohydrate Interactions
Enzymes with Novel Properties
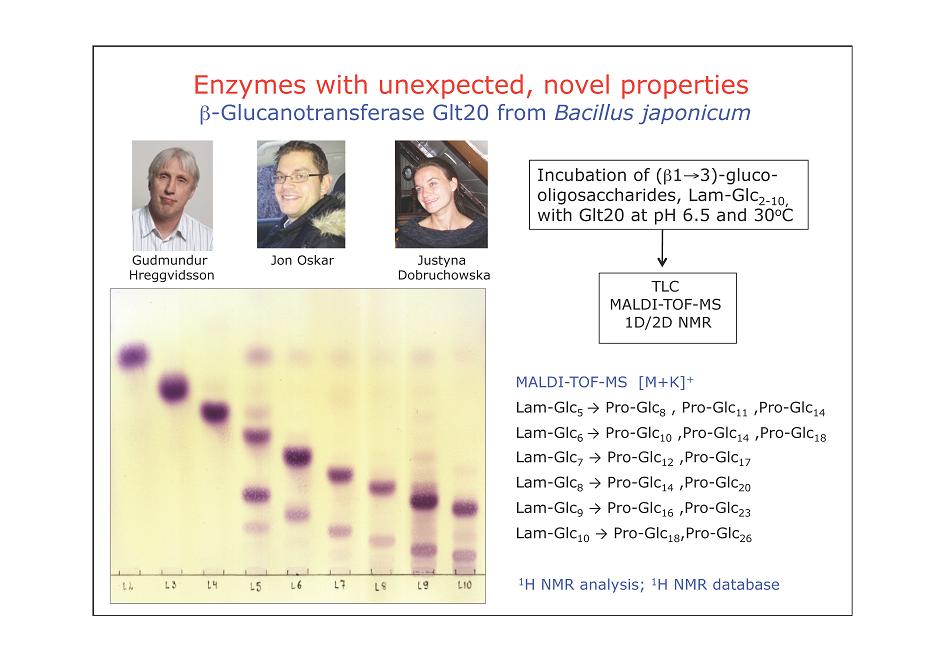
Conclusion
The Author
Hans Kamerling Johannis P. Kamerling (1944) studied chemistry at Utrecht University (1962-1969), and obtained his Ph.D. degree in 1972 with Prof. J.F. Arens and Prof. J.F.G. Vliegenthart. His glycoscience research is focused on (i) structural studies of polysaccharides and glycoprotein glycans (since 1969) ; (ii) organic/enzymatic synthesis of (conjugated) carbohydrates (since 1984) ; and (iii) carbohydrate-mediated interactions […]
Introduction
The elucidation of the 3D structures and dynamics properties of oligosaccharides and glycoconjugates, both in the free state and bound to proteins, is a prerequisite for a better understanding of the molecular basis of their associations and interactions, and the relationships between structures and functions, which are involved in the biochemistry of recognition processes and […]
The Origin of the Nuclear Overhauser Effect
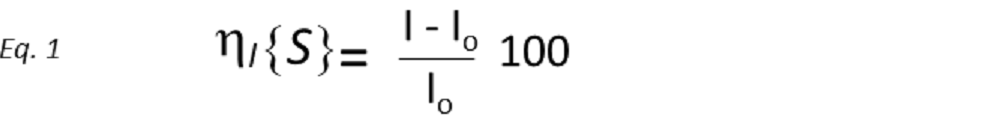
An accurate definition of the Nuclear Overhauser Effect (NOE) is the change in intensity of one resonance when the spin transitions of a dipolarly coupled nucleus are somehow perturbed from their equilibrium populations. This perturbation is achieved by either saturating a resonance, i.e., equalising the spin population differences across the corresponding transitions (then it is […]
Rotating Frame NOE: ROE
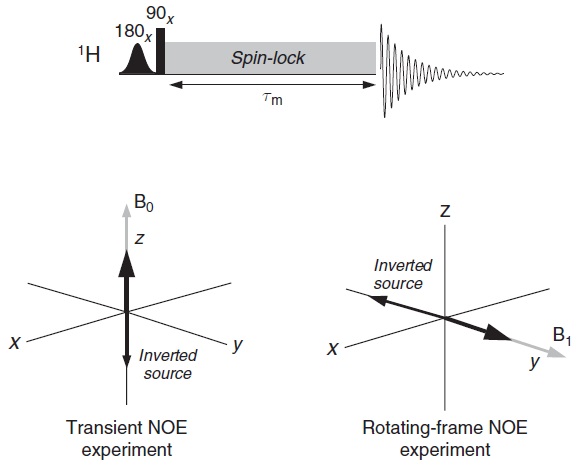
The greatest problem associated with NOE experiments is the zero-crossing region around ω0τc ≈ 1 where the conventional (laboratory-frame) NOE observed via steady-state or transient techniques becomes vanishingly small. This typically occurs for mid-sized molecules with masses of around 1000-2000 Da, depending on solution conditions and spectrometer frequency. With the increasing interest in larger molecules […]
Measuring Internuclear Distances
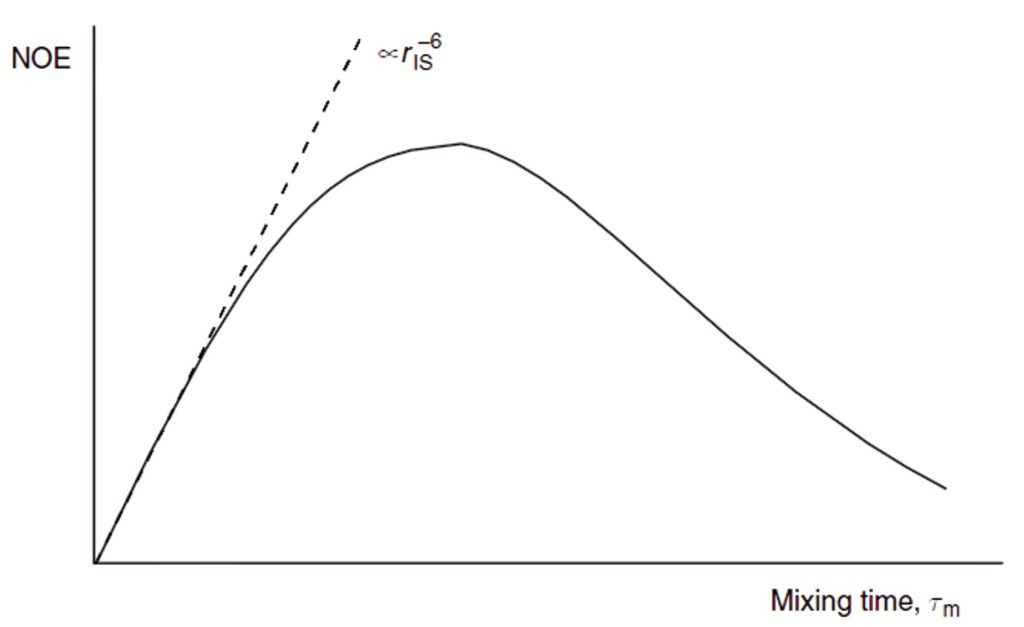
As it has been commented in the previous section, the NOE is the result of the dipolar coupling of two spins which are close in the space to each other, so that this is a distance dependent effect. However, the exact NOE dependency with distance and which NOE experiment we must carried out to obtain […]
The Off-Resonance ROESY Experiments
In an anisotropic molecule, the NOESY or ROESY based distances calculated using the Isolated Spin Pair Approximation using a single distance (and therefore a single correlation time) as reference are not accurate as they depend on the angle between the interprotonic vector and the molecular axis, that governs the correlation time of the vector. As an alternative […]
References
The Authors
Juan Carlos Munoz Garcia After graduating as a Bachelor of Science in Chemistry from the University of Almería (Spain), Juan moved to the University of Malta as a Leonardo Da Vinci intern, where he worked on the extraction and bioactivity determination of plant-derived extracts. In 2009, he joined the Glycosystems Laboratory of the Institute for […]
Introduction
A critical step in drug discovery is the identification of high-affinity ligands for specific macromolecular targets. This is why, over the last 10 years, NMR spectroscopy has become a powerful tool in the pharmaceutical industry. Furthermore, significant improvements have been carried out in the field of NMR instrumentation in recent years, e.g., digital recording, cryogenic […]
The Equilibrium Kinetic of binding study
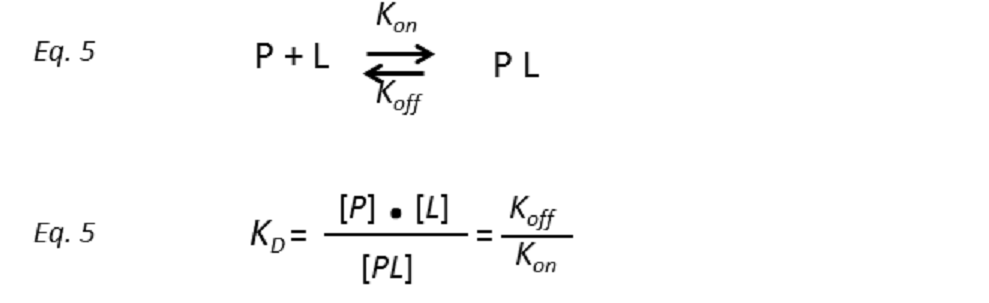
Assuming a protein receptor P with a single binding site for the ligand L, such that they interact to transiently form the complex PL, the binding equilibrium can be described by a model of two states with a temperature dependent dissociation constant KD: where the unimolecular dissociation rate constant koff is inversely related to the […]
Transferred NOESY EXperiments (tr-NOESY).
The theory of transferred NOE has been described in numerous publications (Balaram et al., 1972a) (Balaram et al 1972b) ( Clore & Gronenborn, 1969) (Moseley et al., 1995) and several excellent reviews have been written (Neuhaus & Williamson.,2000) (Ni.,1994) (Post, 2003) . Since the kinetics of the majority of glycan-protein interactions is fast and with […]
Saturation Transfer Difference Spectroscopy (STD).
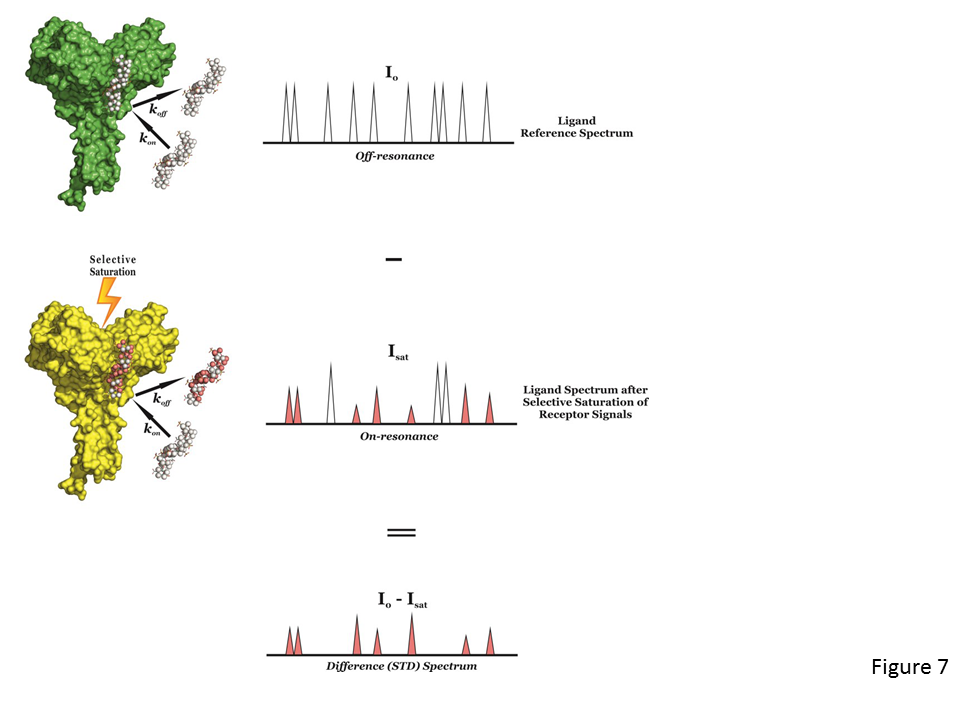
The STD NMR experiment (Mayer & Meyer, 1999) is another spectroscopic technique to study the interactions, in solution, between a large molecule (receptor) and a medium-small sized molecule (ligand), and, alike tr-NOESY, it is based on the Nuclear Overhauser effect and the observation and analysis of the resonances of the ligand protons. The experiment is carried out […]
References
The Authors
Juan Carlos Munoz Garcia After graduating as a Bachelor of Science in Chemistry from the University of Almería (Spain), Juan moved to the University of Malta as a Leonardo Da Vinci intern, where he worked on the extraction and bioactivity determination of plant-derived extracts. In 2009, he joined the Glycosystems Laboratory of the Institute for […]
The mechanism of HIV entry to the host cell
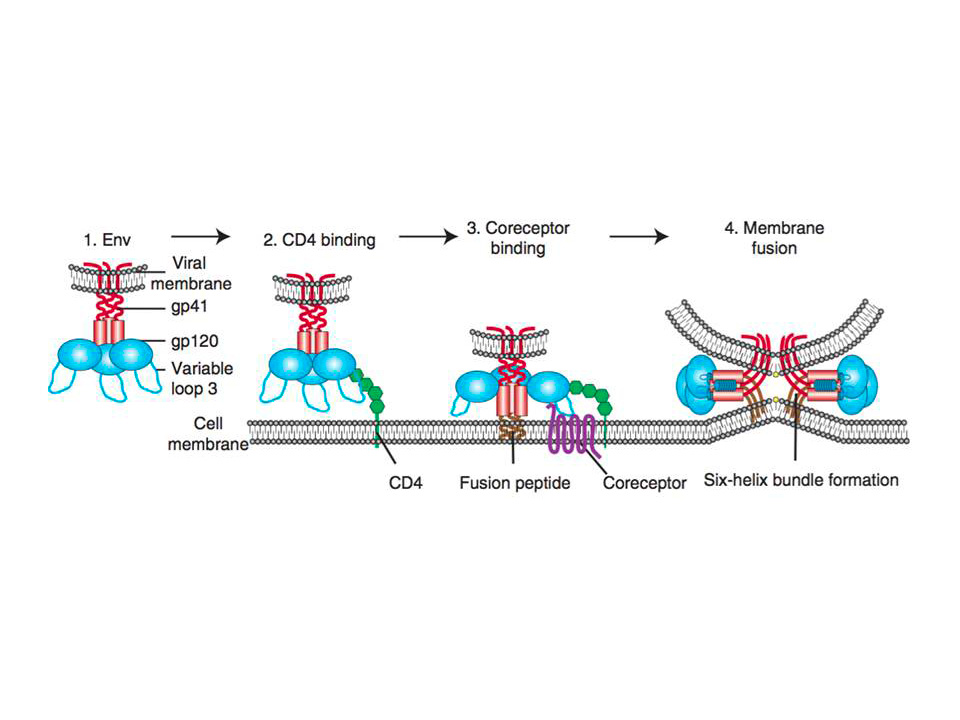
The host cell infection by HIV starts by the entry of the virus into the cell. The major target cells of HIV are CD4+ T lymphocytes, which also express chemokine receptors CCR5 and CXCR4 that are exploited by HIV to enter the cells, hence also called HIV co-receptors. The virus entry starts with the adhesion […]
Dendritic cell implication in HIV-1 sexual transmission
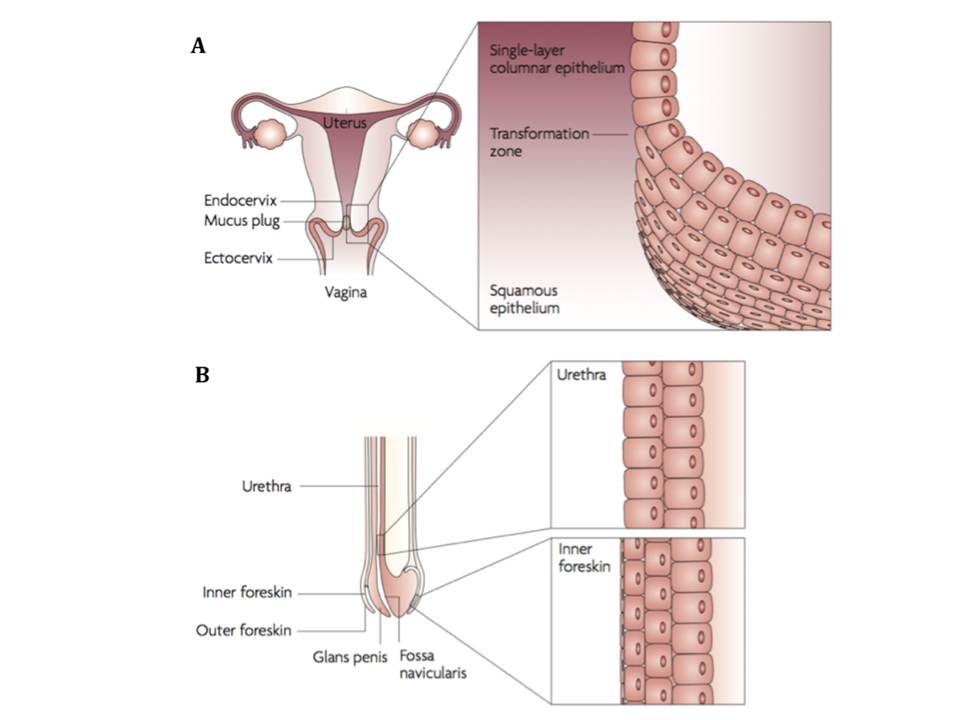
Mechanism of HIV invasion of genital mucosa Sexual transmission through genital and rectal mucosa accounts for the vast majority of HIV- 1 infections worldwide. The viral invasion in women occurs mostly through the non-keratinized squamous epithelium of the vagina and ectocervix, and also through the single-layer columnar epithelium of the endocervix (figure 2A). Sexual transmission […]
Structure and function of C-type lectin receptor DC-SIGN
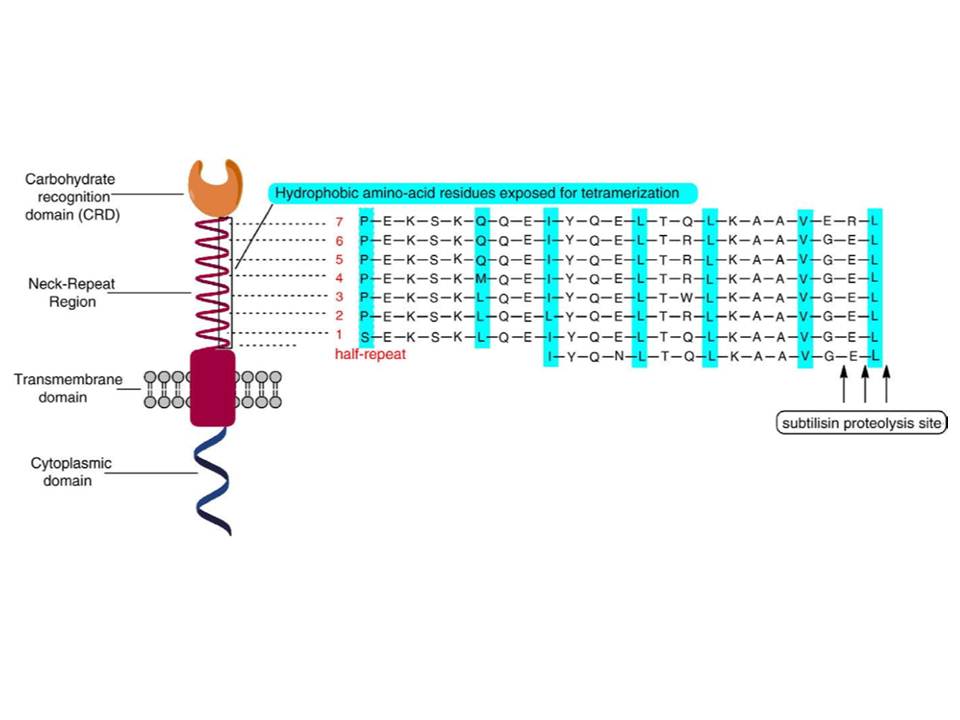
Structure and function of C-type lectin receptor DC-SIGN DC-SIGN is a type II membrane protein with a single CRD, i.e. a type II C-type lectin. It was first cloned and identified as a C-type lectin binding to HIV glycoprotein gp120 (Curtis, B. M, Scharnowske, S, &Watson, A. J. 1992) and later rediscovered and renamed (Geijtenbeek, T. B, Torensma, […]
Langerin: a specific CLR of Langerhans cells
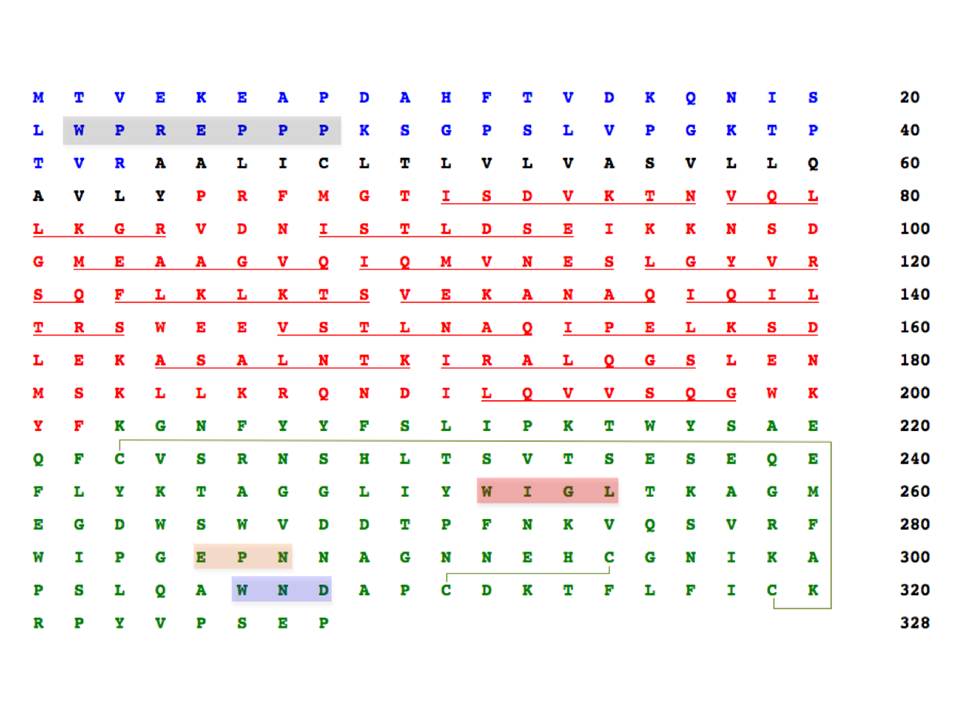
As DC-SIGN, langerin is a type II C-type lectin, almost exclusively expressed on epidermal LCs, but also present on dermal CD103+ DCs and lymph node resident CD8+ DCs (Valladeau, J, Ravel, O, Dezutter-Dambuyant, C, et al., 2000, Romani, N, Clausen, B. E, & Stoitzner, P. 2010, doyaga, J, Suda, N, Suda, K, et al., 2009). LCs is […]
The comparison of the CRDs of DC-SIGN and langerin
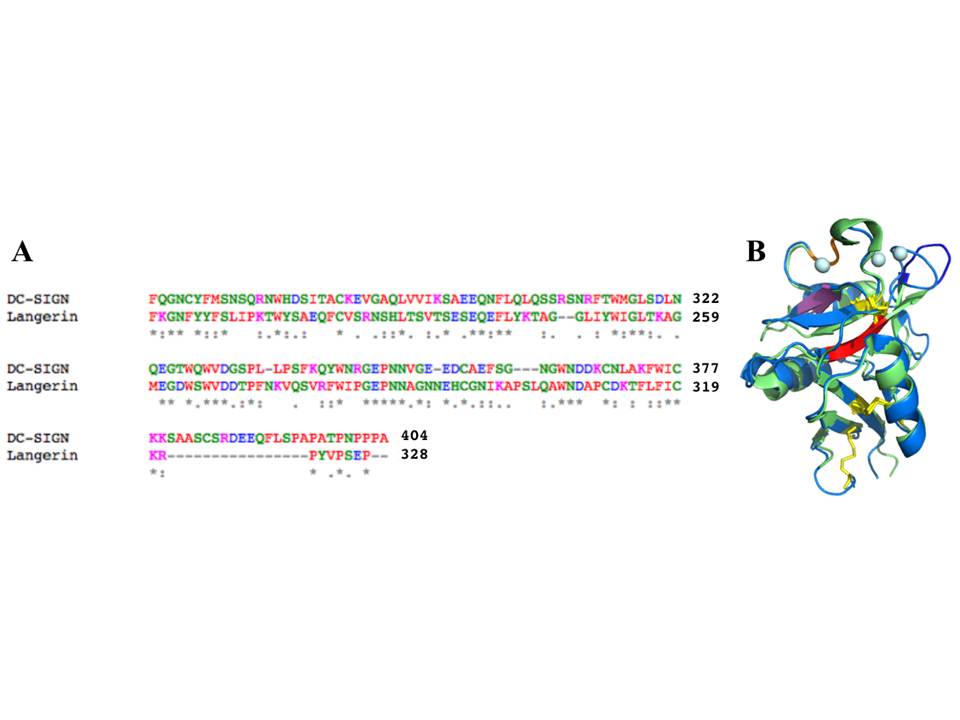
Both DC-SIGN and langerin are lectins of type II with C-type CRDs. The sequences and the overall structures of their CRDs are close (fig. 22) and correspond to the classical CRD fold However, several important differences can be tracked in these two CRDs. Langerin has several structural elements that are not present in DC-SIGN CRD, […]
References
Arthos, J, Cicala, C, Martinelli, E, Macleod, K, Ryk, D. V, Wei, D, Xiao, Z, Veenstra, T. D,Conrad, T. P, Lempicki, R. A, Mclaughlin, S, Pascuccio, M, Gopaul, R, Mcnally, J, Cruz, C. C, Censoplano, N, Chung, E, Reitano, K. N, Kottilil, S, Goode, D. J, & Fauci, A. S. (2008) Hiv-1 envelope protein binds […]
Authors

After graduating as a Master of Science (Biochemistry, Molecular Biology, and Biopharmacy) from the Vilnius Gediminas Technical University, Ieva spent three years as an analyst working within the company : Sicor Biotech – TEVA pharmaceuticals, in Vilnius. There she carried on the evaluation of biopharmaceutical interactions with their receptors using Surface Plasmon Resonance. In 2009 […]