
The immune system has evolved to an amazing defense mechanism against the invasion of a broad variety of pathogens and parasites. It is possessed by the majority of the living organisms including such simple beings as unicellular bacteria or invertebrates, where it exists in a rudimental form (He, J & Deem, M. W. 2010, Rowley, A. F & Powell, A. 2007). In higher vertebrates, including humans, it has evolved to an extremely sophisticated system that has the features of specificity and memory, and which is implemented by the two components – innate and adaptive immunities. The adaptive immunity is highly specific with a unique property of memory and it requires stimulation with an antigen prior to the response, whereas, innate immunity is less specific and has no memory. However, innate immunity has a crucial role (fig. 1), as it is the first line defense of the organism against the invading pathogens and its responses activate the adaptive immunity. The importance of innate immunity is also highlighted by the fact that the defects of its components are relatively rare and almost always lethal (Goldsby, R, Kindt, T, Osborne, B, & Kuby, J. 2002, Parham, P. 2009)
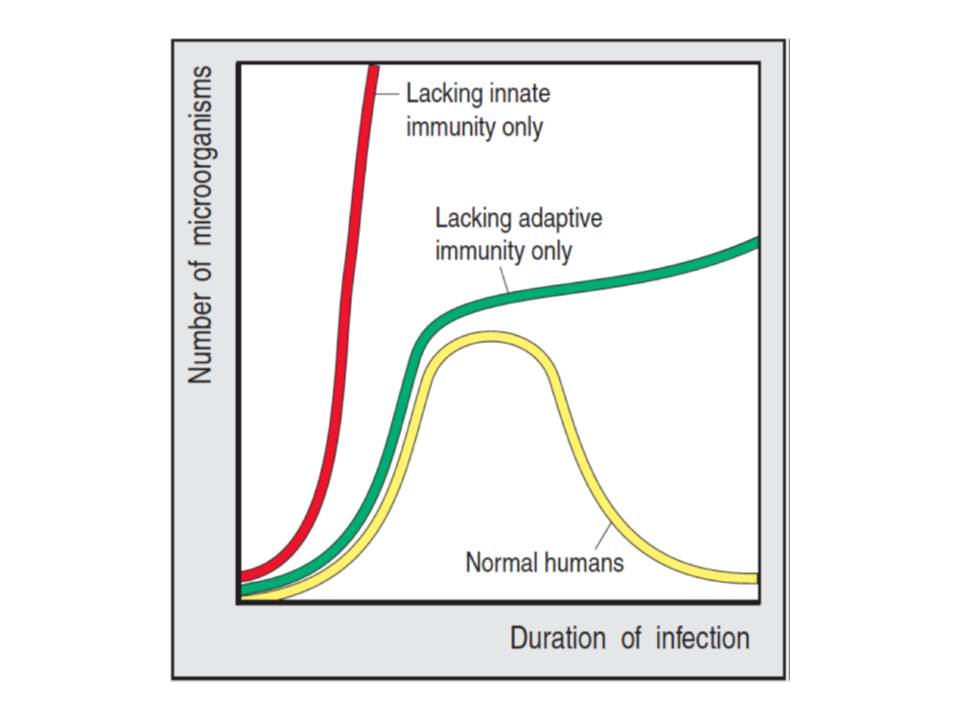
The innate immunity comprises several levels : anatomic, chemical-physiological (temperature, pH, soluble factors), cellular, and inflammatory barriers (fig. 2), which are closely related and work together to provide the effective protection.
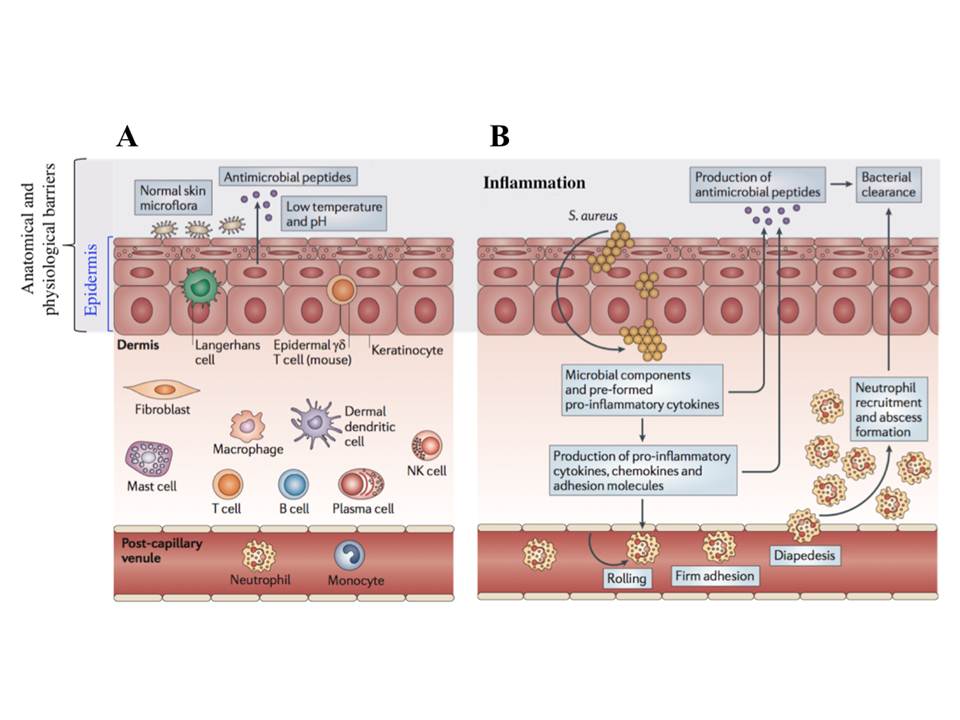
The anatomical barrier includes the skin and mucosa, and provides a physical obstacle for the pathogen invasion. Its importance is obvious as in the case of the loss of the integrity of the body’s internal epithelia, infection is a major cause of mortality and morbidity (Murphy, K, Geha, R, & Notarangelo, L. 2011).