1893. Defining Carbohydrates.
“It appears, therefore, that we have an homologous series of aldehyde or ketone alcohols of the general formula CnH2nOn with these common properties:
- 1. Sweet to the taste ;
- 2. Optically active ;
- 3. Reducing alkaline metallic solutions ;
- 4. Yielding with phenylhydrazin characteristic crystallinic compounds.
- 5. Other reactions, such as great solubility, decomposition by heat, and color reactions, are less characteristic, although possessed in common. “
With this list, W. E. Stone described the use of the term “Carbohydrates” in 1893. w.e. stone, 1893 The term “carbohydrates” was well defined and explained to the scientific community: w.e
“Those containing three or multiples of three, carbon atoms undergo alcoholic fermentation with yeast, and this periodical reaction seems an additional argument for their common nature. Individually they yield, when heated with strong acids, characteristic derivatives; for instance, the pentoses yield furfurol; the hexoses levulinic acid; others have not been carefully studied in this direction… E. Fischer proposes to apply the name “sugars” to all the members of this homologous series, to which he has lately added the glycol-aldehyde C2H4O2 as the simplest possible example. The popular conception of the properties of a sugar are not, however, easily reconciled with the properties of some of these bodies, while “carbohydrates” at least possess some reference to their impirical composition.”
1931. Trigerring the adaptative immune system.
One the most studied behavior of carbohydrates (especially bacterial capsular polysaccharides) is linked to their ability in triggering the adaptive immune system to produce protective antibodies. Avery and Goebel described one of the seminal works on conjugate carbohydrate-vaccines in 1931 o.t. avery & w.f. goebel, 1931
“Type-specific antipneumococcus immunity has been induced in rabbits by immunization with antigen prepared by combining a specific derivative of the capsular polysaccharide of type III Pneumococcus with globulin from horse serum.”
The field of carbohydrate-based vaccines is still exploiting the same 1931 strategy in designing synthetic and semi-synthetic vaccines with successful examples against different bacteria.
1931. Colloidal particles as carriers for carbohydrate-based vaccines.
The same year of the Avery and Goebel publication, organic colloidal particles were explored by Zozaya as carrier for bacterial polysaccharides trying to improve their immunogenic properties. The adsorption of polysaccharides to colloid surface was tried to have an “adjuvant-effect”. The work of Zozaya showed for the first time that polysaccharides can be rendered immunogenic by adsorption on colloid carriers. j. zozaya, 1931
“Polysaccharides, as such, when injected have not proven to result in antibody formation. Many have assumed that fairly complex nitrogenous substance had to be a part of the polysaccharide combination to make these antigenic. It occurred to us, however, that this might not be necessary and that combinations might be prepared which may owe their antigenic properties to a physical basis or arrangement rather than to an organic chemical combination.(I am aware of Langmuir’s idea of the chemical nature of adsorption in colloids.)”.
At that time, the word “nanotechnology” was “uncommon” but the approach of Zozaya is still explored in designing new nano carriers for carbohydrates-based vaccines.
1965. Serological relationships of bacterial polysaccharides.
The interest in investigating the immunological properties of bacterial polysaccharides led Shifrine and Gourlay to describe in 1965 the serological relationship between Galactans from normal bovine lung and from Mycoplasma mycoides m.shifrine & r.n. gourlay, 1965. A pneumogalactan isolated from bovine lung was able to precipitate pneumococcal polysaccharide type XIV antiserum suggesting that both polysaccharides have galactose residues of similar linkages. The cross-reactivity of pneumogalactan and the galactan from Mycoplasma mycoides indicated that both polysaccharides have some common linkages. The authors also described the possibility that pneumogalactan plays a role in the bacterial pathogenesis either in allergic type reactions. The correlation between specific immunological patterns and specific immunological responses was than elegantly proven. m.shifrine
Little by little, the first lectins were explored as a biochemical tool to recognize carbohydrates. The immunological responses in vivo were also investigated with lectins from a serological point of view.
1967. The discovery of IgG glycoforms.
Leon showed for the first time the different IgG glycoforms present on the cancer serum analyzing the ConA reactivity of forty-two IgG myeloma sera. m.a. leon, 1967 m.a.
“If one accepts the theory that monoclonal myeloma protein in an individual serum is homogenous with respect to amino acid sequence, then all the molecules of myeloma protein have the same range of conformational possibilities in equilibrium. Carbohydrate chains attached to these molecules should show similar reactivity with ConA provided that the carbohydrate chains are identical and are attached at the same position in the peptide chain. The partial precipitation of IgG myeloma proteins by ConA, demonstrates existence of some type of heterogeneity of each IgG myeloma carbohydrate. The fact that not all of the protein was precipitated and that the amounts precipitated varied from serum to serum may he interpreted as demonstrating heterogeneity of the carbohydrate in these myelonma proteins.”
IgG isoforms are nowadays a critical issue in the biopharmaceutical world due to their different properties and efficacy in vivo.
1969. Role of protein-glycoforms in transplantation.
The study of different protein-glycoforms was also applied in the field of transplantation. In 1969 the molecular and chemical properties of membrane components bearing the major transplantation antigen system in mouse (H-2) and humans (HL-A) were compared and found to be extremely similar except for their glucosamine content. d.l. & s.g. nathenson, 1969 d.l.
“These physical and chemical similarities in the characteristics of molecules bearing the transplantation antigens are in accord with biologic studies indicating a comparable functional immunologic role for the mouse H-2 and human HL-A antigen systems. The greatest difference was seen in the glucosamine content, which was less than 0.05 per cent in the HL-A materials and in the order of 5 per cent in H-2 bearing material. Neutral sugar composition of the alloantigenic material from these two animal species show somewhat greater similarity and all samples had about 1 per cent sialic acid.”
1970. Carbohydrates modulate the behavior of proteins.
The critical role of carbohydrates in modulating the behavior of proteins was clear and in 1970 the attachment of carbohydrates to the variable region of myeloma immunoglobulin light chains was demonstrated and analyzed (see Figure 1). h.c. sox jr. & l. hoodt, 1970
 h.c.
h.c.
The function of carbohydrate in immunoglobulins is unknown although the remarkable conservation of heavy chain carbohydrate throughout mammalian evolution suggests an important role for carbohydrate attached to heavy chains. The postulated role for carbohydrate in facilitating secretion of immunoglobulin from the cell could be served by the oligosaccharide present on all heavy chains. The function of variable region carbohydrate is likewise unknown. It seems unlikely that carbohydrate plays an important role in antigen binding since the oligosaccharide is a bulky group made up of a limited variety of carbohydrate residues and cannot be synthesized in a precisely reproducible fashion. In fact, the bulky carbohydrate group attached near the antigen-binding site could interfere with antigen binding. Were this the case, one would predict that the variable regions of light chains from normal serum immunoglobulins should have little or no carbohydrate bound near the antigen-binding site, since interaction of antigen with antibody bound to the surface membrane of immunocyte precursors is apparently necessary for induction of the immune response. This possibility is currently being investigated.
The fascinating conclusions of the authors are still “modern”. The role of Fc glycosylation was extremely studied in the last 30 years and recently also the variable chains of antibodies have attracted again glycoscientists to investigative their role in immune-recognition as proposed in 1970.
1970. Lectins are named.
Lectins were one the best tools to interrogate biological systems on their carbohydrates repertoire. The blood group specific plant agglutinins (named lectins by Boyd) were discovered in 1945 by Landsteiner and reviewed by Boyd in 1970. w.c. boyd, 1970
w.c.

“One day toward the end of 1945, looking at this table in the second English edition of Landsteiner’s book, I was seized with the idea that if such extracts could show species specificity, they might even show individual specificity; that is, they might possibly affect the red cells of some individuals of a species and not affect others of the same species. Therefore, I asked one of my assistants to go out to the comer grocery store and buy some dried lima beans. Why I said lima beans instead of the more common pea beans or kidney beans I shall never know. But if we had bought practically any other bean we would not have discovered anything new.”
Boyd proposed that these blood-antigen-specific plant agglutinins were called “lectins” from the Latin legere, to pick out or choose. 27 different lectins were listed in this review with their activities and source. At that time the main important activity of lectins was related to their specificity in agglutinate specific blood antigens.

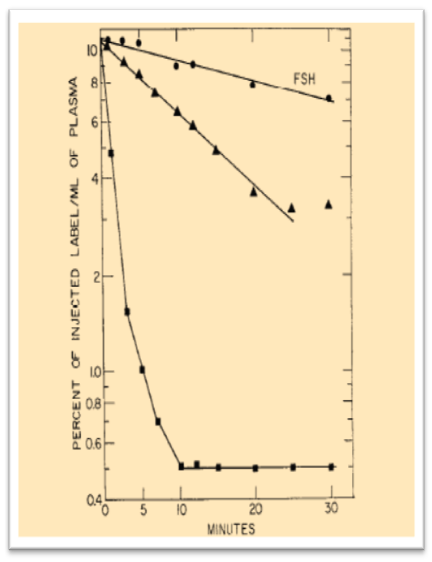
1971. Role of Sialic acid.
More biochemical tools were exploited to investigate the biological role of carbohydrates on proteins-fate in vivo and in 1971 the role of sialic acid in determining the survival of glycoproteins in the circulation was demonstrated. a.g.morell et al., 1971 Evidence is presented to indicate a generalized role for the terminal sialic acid residues of circulating glycoproteins. Upon injection into rats, all of the desialylated plasma proteins tested, with the exception of transferrin, were promptly removed from the circulation and were recovered from the liver. Figure 4 shows the plasma survival time of native and desialylated follicle-stimulating hormone in rat.
1971. Assessing the importance of a proper and conserved protein glycosylation.
The importance of a proper and conserved protein glycosylation was little by little becoming a dogma in the field and the glyco-code was becoming something difficult to decipher especially when glycoscientists investigated that code among different species. A nice example of the co-evolution of the glyco-cocde among different species was described in 1971. j.a. clegg & s.r. smithers, 1971 The parasitic worm Schistosoma mansoni may live in the portal circulation of humans for many years. In rhesus monkey, established adult worms persist even though their presence stimulates powerful host immunity against reinfection. It was suggested that this concomitant immunity could be explained if the adult worms possess host antigens to evade host immune responses. The existence of these host antigens at the surface of the worms (carbohydrates) was demonstrated by transferring worms (grown to maturity in mice) directly into the portal circulation of rhesus monkeys immunized against mouse erythrocytes.
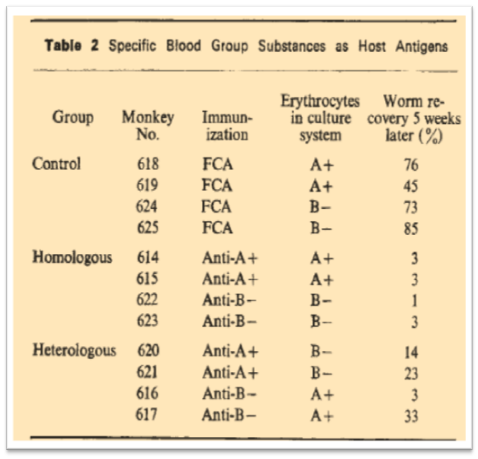
When the transfer was homologous i.e. A+ worms into anti-A+ monkeys and B- worms into anti-B- monkeys, almost all Schistosomula were destroyed. With heterologous transfer, i.e. A+ worms into anti-B- monkeys and vice versa, survival rates were more variable. The authors concluded that the schistosomula of S. mansoni can acquire human antigens and that these antigens can be acquired during cultivation in vitro. Nowadays we know that this parasitic worm has its own glycosylation machinery but the observations of Clegg and Smithers were extremely important.
1972: Lectins as carbohydrate-binding proteins.
In 1972 an impressive review about lectins was published resuming all the preliminary knowledge on these carbohydrate-binding macromolecules. n. sharon & h. lis, 1972 n.
“Proteins that possess the remarkable ability to agglutinate erythrocytes and other types of cell are widely distributed in nature. Such agglutinins are found predominantly in the seeds of plants, in particular those of the legumes, but they are also present in other parts of plants such as roots, leaves, and bark. In addition, they occur in invertebrates, for example, in snails and the horseshoe crab, and in lower vertebrates such as fish. Plant agglutinins are commonly referred to as phytohemagglutinins or phytoagglutinins. However, as cell-agglutinating proteins also occur in organisms other than plants, the term “lectins,” proposed by Boyd, appears more suitable and will be used in this article.”
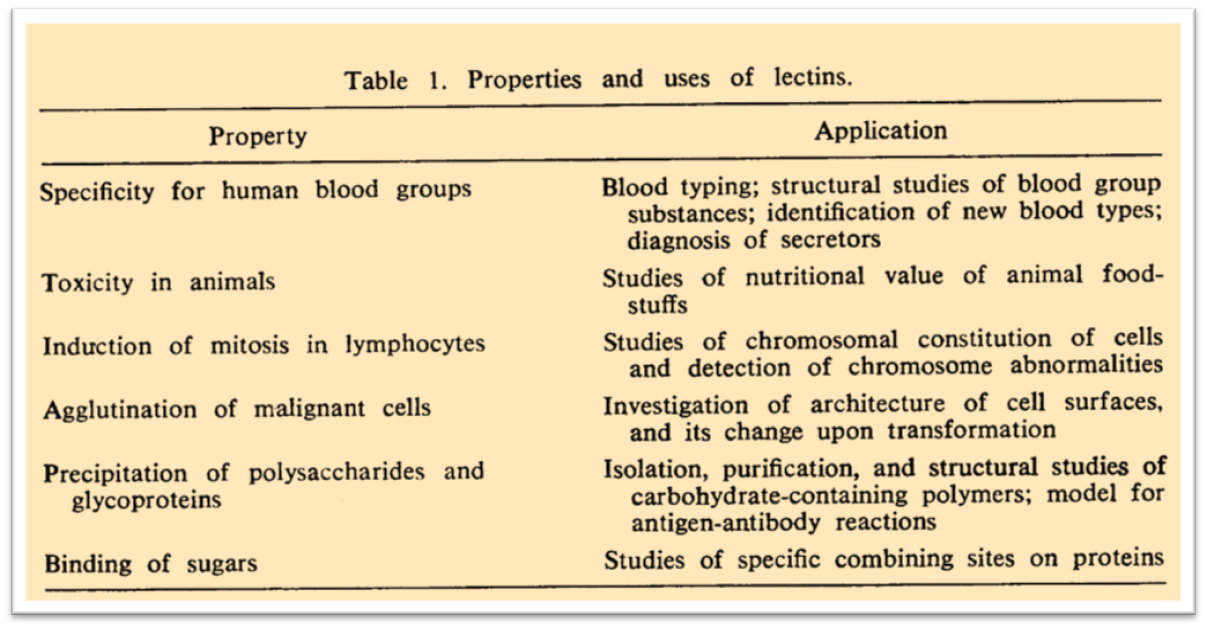
In this review one of the first tables with the known lectins properties was published. The specificity for defined carbohydrates and their source was listed in the Figure below:
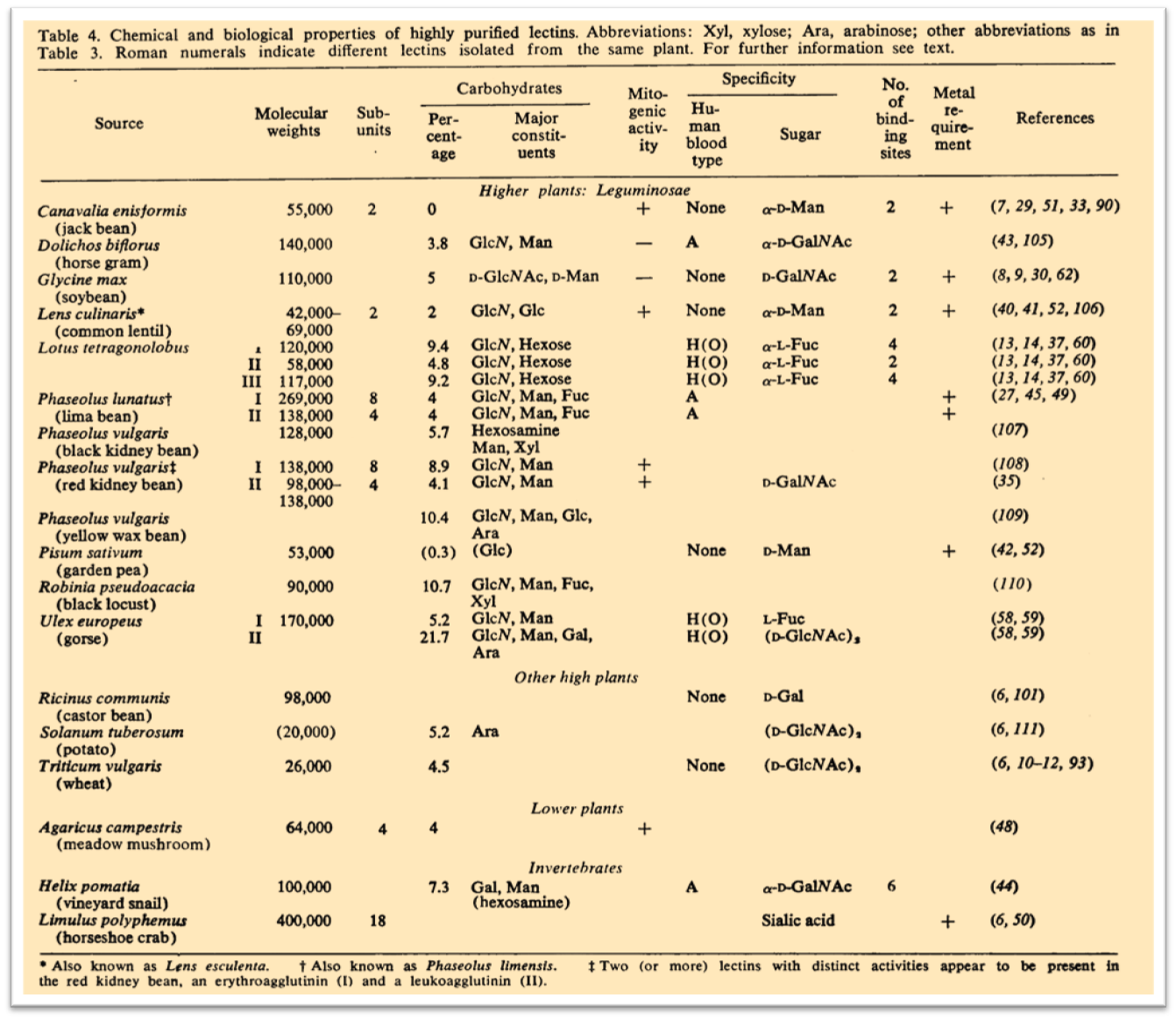
The conclusive sentence of this review is really fascinating:
“The role of lectins in nature remains a mystery. It has been suggested that they are antibodies intended to counteract soil bacteria; that they serve to protect plants against fungal attack by inhibiting fungal polysaccharases; that because of their affinity for saccharides they are involved in sugar transport and storage; or that they serve for the attachment of glycoprotein enzymes in organized multienzyme systems. In the last 30 years we have learn the importance of lectins in different fields of immunology and we do not think of lectins as simple tool to precipitate or agglutinate carbohydrate-containing macromolecules.”
Notwithstanding the pioneer works of different glycoscientists with lectins were of a key importance in understanding biological systems and their interactions with carbohydrates.
1974. First analysis of a cancer cell glycocalix.
In 1974 the alterations of membrane glycopeptides in colonic adenocarcinoma were studied. y.s. kim et al., 1974 The examination of blood group activity and the lectin-binding properties of these membrane glycopeptides revealed the changes of specific carbohydrate structures in the tumor tissue. A cancer cells glycocalix composition was analyzed for the first time as described in Figure 8
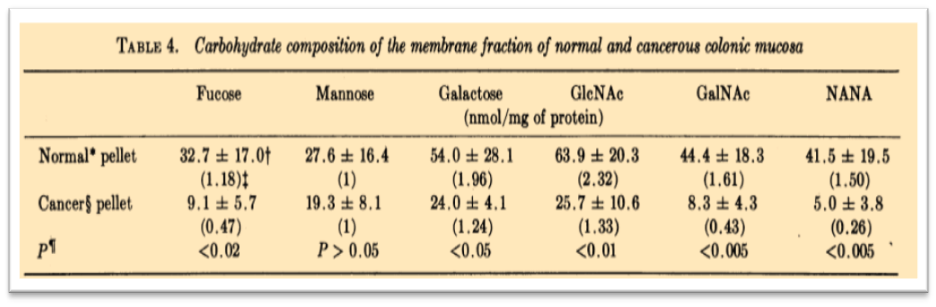
The results from this work proved the alteration in glycoprotein biosynthesis during tumorigenesis:
“whether the alterations in the biosynthesis of glycoproteins and glycolipids are primary events or reflect changes in the tissues secondary to neoplastic transformation remains to be determined, but these changes seem to be a common manifestation of the malignant process”.
1975. Existence of “natural” anti-carbohydrate antibodies.
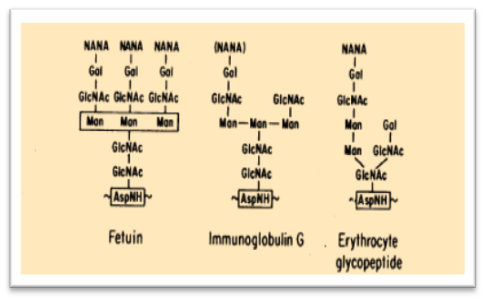
In 1975 with a series of precise and elegant experiments, the existence of “natural” anti-carbohydrate antibodies was demonstrated and characterized for different species, concluding with the concept of animal tolerance to “self” carbohydrate-antigens.b.a. sela et al., 1975
“The origin of natural antibodies to carbohydrate structures is not known. It has been noted that when animals are immunized with a glycoprotein such as fetuin, the elicited immune response is predominantly directed at the protein portion of the molecule and not at the carbohydrate moiety. The difficulty of raising antibodies to sugar portions of glycoproteins may be due to the obvious similarities (Figure 9) in many of the carbohydrate structures appearing in otherwise unrelated serum glycoproteins such as fetuin, thyroglobulin, Ig molecules of different classes, or cell surface glycopeptides, including the histocompatibility antigens. The similarities of the carbohydrates may have rendered the animals tolerant to these structures, which would be recognizable as “self” antigens.”
1976. The first review on gangliosides appears in literature.
In 1976 one of the first review on gangliosides appears in literature. The biosynthesis and function of these natural glycoconjugates were described unifying all the previous observations in the field. p.h. fishman & r.o. brady, 1976
“Gangliosides comprise a family of acidic glycolipids that are characterized by the presence of sialic acid. They are unusual compounds in that they contain both hydrophilic and hydrophobic regions, and they bear a strong negative charge. Gangliosides are primarily membrane components, and plasma cell membranes are highly enriched in these materials.”
An impressive “hypothetical” mechanism of action between cholera toxin and human GM1 (monosialotetrahexosylganglioside, the “prototype” ganglioside) was proposed (see Figure 10) including also a preliminary concept of carbohydrate-multivalency during the viral attachment on human cells.
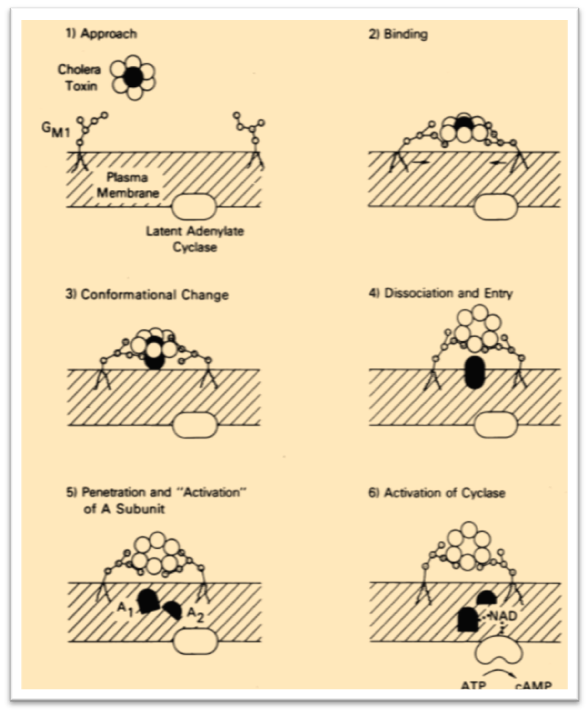
1978. The antigenicity of surface carbohydrates is described.
In 1978 the importance of surface carbohydrates on the antigenicity of two different viral glycoproteins was described.l.j. van eldik & al., 1978 The antigenicity of one of them (the gp85 from Avian Myeloblastosis Virus, AMV) was decreased after glycosidases treatment. l.j.
“These results indicate that the immunological reactivity of AMV gp85 depends upon an intact carbohydrate side chain. Digestion of AMV gp85 with proteases and glycosidases showed that protease treatment resulted in a partial loss of antigenic reactivity. The proposed structure for the carbohydrates on the surface of the AMV gp85 was proposed clearly showing a similarity with the structure of many serum glycoproteins.”
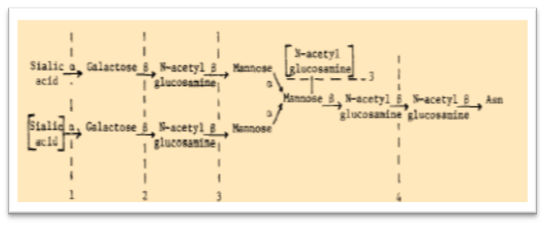
1978. Description of the different IGg subclasses of anti-carbohydrate antibodies.
A study on the different IgG subclasses of anti-carbohydrate antibodies was described.r.m. perlmutter et al., 1978 In that paper the authors reported that mouse IgG antibodies directed against groups A and C streptococci, α-(1→3) dextrans, and phosphocholine are mainly restricted to mouse IgG3 subclass.
” Recruitment of the rare IgG3 subclass by these antigens suggests that this constant region may possess unique biologic properties perhaps relevant to bacterial immunity. Interestingly, mice are not the sole species in which carbohydrate antigens appear to elicit an IgG response restricted to a single subclass. Human antibodies to carbohydrates have been reported to be restricted mainly to the human IgG2 subclass.”
This described recruitment of a minor immunoglobulin isotype by carbohydrate antigens supports the concept of immune-regulation at the level of subclass, suggesting that mammals may share a structurally similar isotype with a common evolutionary origin.
1979. Gangliosides and immunosuppression in cancer
In 1979 it was described that Con A-treated thymocyte cultures in medium with exogenous gangliosides had a lower rate of thymidine and uridine incorporation into nucleic acids.e.e. lengle et al., 1979
” It was tempting to suggest that gangliosides shed by tumor cells could be the cause of the generalized immunosuppression that is usually observed by decreased responses to mitogen and delayed hypersensitivity reactions…There is substantial evidence that immunosuppressive factors present in sera of tumor-beaming humans and animals are capable of inhibiting transformation of normal homologous lymphocytes. Presumably, such factors either are elaborated directly by the neoplastic cells or are produced by normal cells in response to the presence of the tumor. The recent observation that elevated serum levels of glycohipids are found in both human and mumine tumor bearers suggested an investigation of ganghiosides as immunosuppressive agents. Furthermome, artificial elevation of membrane glycolipid cornposition by addition of exogenous glycolipids reduced the growth rate of both normal and transformed cell lines.”
1981. Altering the glyco immune-synapse by mean of monosaccharides
In 1981 the interactions between regulatory T lymphocytes and other cells were studied under the influence of several monosaccharides. u.h koszinowski & m. kramer, 1981 A first hypothesis for a glyco immuno-synapse existence between these type of cells was proposed: Carbohydrates on cell membrane-inserted glycoproteins or glycolipids might function as specific ligands for recognition by cellular receptors or soluble factors. To disrupt and affect this “general” glyco immune-synapse, it was reported that the addition of monosaccharides can counteract the effect of T suppressor cells during the generation of alloreactive cytotoxic T cells in vitro and the most prominent effect was described for methyl-α-D-mannopyranoside. The authors suggested the hypothesis that recognition structures on T suppressor cells (or secreted T suppressor cells molecules), mediate the immune-suppression interacting with complementary structures on their target cells, and the studied monosaccharaides added to the cells competitively blocks these interactions. u.h
1984. ELLA: An Enzyme-Linked Lectin Assay is described.
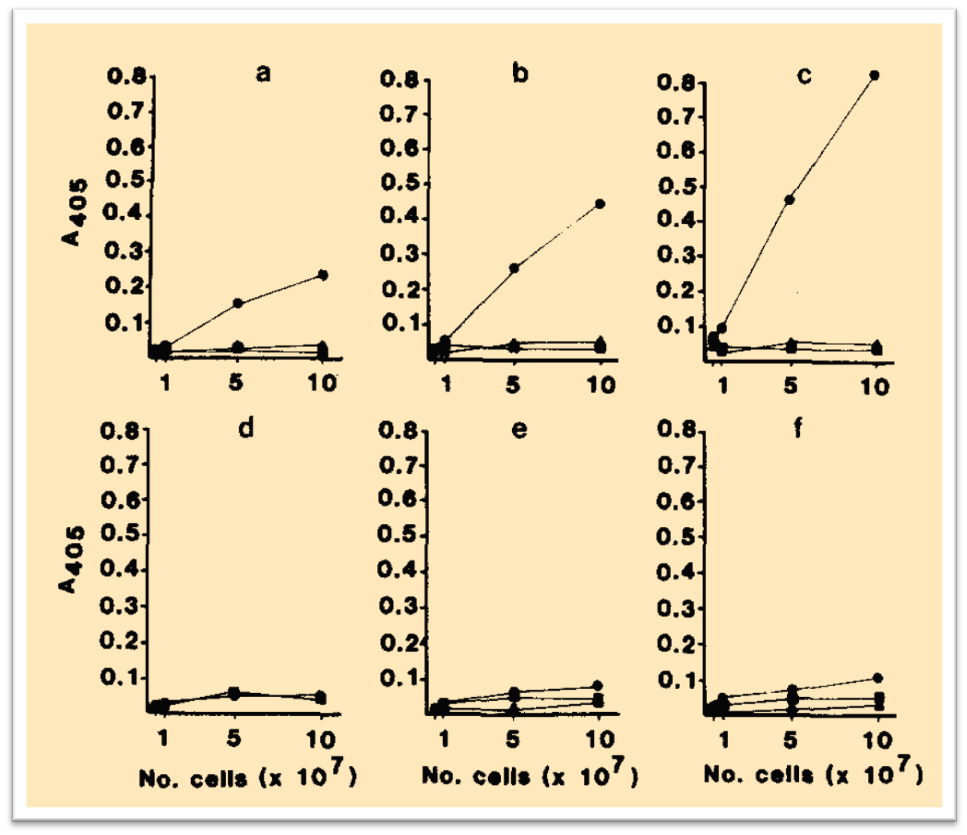
One of the first solid-phase carbohydrate array was proposed in 1984: an enzyme-linked lectin assay (ELLA) was developed to detect specific carbohydrate units on the surface of unfixed cells.j.p. mccoy et al., 1984
“The conjugation of Griffonia simplicifolia I-B4 (AP-GS I-B4, an α Gal-p binding lectin) to alkaline phosphatase allowed the ELISA-type procedure for the detection of specific carbohydrate-antiogens. The assay may be read in standard ELISA plate readers, since the cell-bound enzyme-lectin conjugate is specifically eluted from the cells prior to development of the conjugate. ELLA, when read in an enzyme-linked immunosorbent assay (ELISA) plate reader, allows better detection and relative quantitation of specific surface carbohydrate units than is possible by standard immunofluorescence with fluorescein-conjugated lectins.”
As described in Figure 12 the AP-GS I-B4 showed a time-dependent affinity for the type-B human erythrocytes. The specificity of the assay was proven with non-reactivity against type-O or type-A erythrocytes.